Гипотеза «бюрократического потолка» для верхней границы сложности генома выглядит особенно правдоподобно в свете почти полного отсутствия роста числа генов в геномах позвоночных, особенно млекопитающих (и в наших геномах тоже), для которых связь между числом генов и размером генома очевидно нарушена (см. гл. 3 и 8). В принципе число генов могло бы быть напрямую ограничено ценой репликации ДНК, но для огромных геномов позвоночных этот фактор можно смело исключить как главное ограничение. Соответственно, цена регуляции, возможно совместно с ценой экспрессии, выглядит наиболее вероятным кандидатом на роль основного фактора, ограничивающего рост числа генов. Поэтому не случайно позвоночные (и в меньшей степени другие многоклеточные эукариоты) выработали новые, замысловатые способы увеличения сложности протеома, такие как широко распространенный альтернативный сплайсинг, альтернативная трансляция и сложная регуляция (в особенности обширный, все еще слабо изученный набор регуляторных РНК). Такие формы сложности не вызывают инфляционный рост числа белок-кодирующих генов и, таким образом, снижают по крайней мере некоторые издержки, особенно издержки трансляции (см. гл. 8).
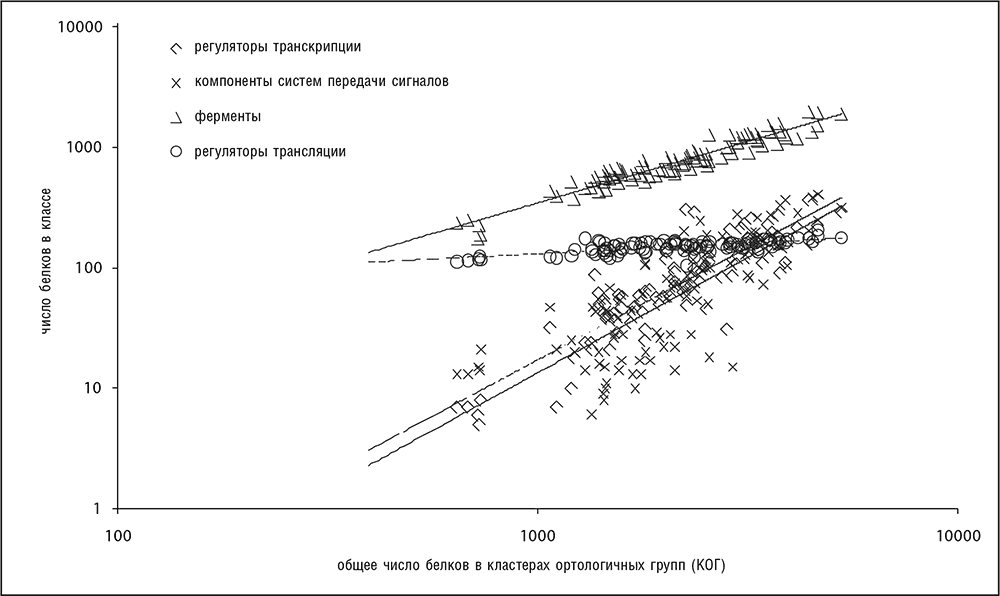 Рис. 4-10. Четко различающиеся соотношения между различными функциональными классами генов и размером генома (общим числом белок-кодирующих генов) прокариот. Использована информация из базы данных COG (Tatusov et al., 2003). График построен в двойных логарифмических координатах.
Рис. 4-10. Четко различающиеся соотношения между различными функциональными классами генов и размером генома (общим числом белок-кодирующих генов) прокариот. Использована информация из базы данных COG (Tatusov et al., 2003). График построен в двойных логарифмических координатах.
Универсальная зависимость численности функциональных классов генов от размера генома находится в обратной зависимости с ранее описанным степенным законом распределения численности семейств генов. Чем больше положительный показатель степени функции зависимости от размера генома для функционального класса генов (рис. 4-10), тем меньше отрицательный показатель степени функции распределения численности семейств в этом классе (рис. 4–7). Кажется интуитивно правдоподобным, что функциональные классы с сильной зависимостью от размера генома должны содержать много больших семейств паралогичных генов. Обратная зависимость этих двух универсалий генома была выведена в рамках простой эволюционной модели, в которой используются правила пропорциональных вычислений для функциональной композиции генома, например «добавить два регулятора на каждый метаболический фермент» (Grilli et al., 2011). Предсказания этой модели подтверждаются эмпирическими данными для многих геномов бактерий и архей.
Стохастичность, нейтральность и отбор в эволюции
Краткий обзор и перспектива: о природе эволюционного процесса
Рекомендуемая дополнительная литература[46]
Глава 5. Сетевая геномика мира прокариот: вертикальные и горизонтальные потоки генов, мобиломы и динамика пангеномов
Размер и общая организация бактериальных и архейных геномов
Пространство-время прокариот и его эволюция
Универсальная зависимость численности функциональных классов генов от размера генома находится в обратной зависимости с ранее описанным степенным законом распределения численности семейств генов. Чем больше положительный показатель степени функции зависимости от размера генома для функционального класса генов (рис. 4-10), тем меньше отрицательный показатель степени функции распределения численности семейств в этом классе (рис. 4–7). Кажется интуитивно правдоподобным, что функциональные классы с сильной зависимостью от размера генома должны содержать много больших семейств паралогичных генов. Обратная зависимость этих двух универсалий генома была выведена в рамках простой эволюционной модели, в которой используются правила пропорциональных вычислений для функциональной композиции генома, например «добавить два регулятора на каждый метаболический фермент» (Grilli et al., 2011). Предсказания этой модели подтверждаются эмпирическими данными для многих геномов бактерий и архей.
Стохастичность, нейтральность и отбор в эволюции
В предыдущих разделах этой главы мы ознакомились со многими количественными универсалиями, отражающими важнейшие аспекты эволюции и функционирования генома. Среди этих универсалий мы рассматривали вездесущий степенной закон распределения, который описывает как структуру всех биологических сетей, так и семейства паралогичных генов в разнообразных геномах, близкое к логарифмически нормальному распределение скоростей эволюции генов и универсальные корреляции, такие как отрицательная корреляция между генной экспрессией и скоростью эволюции. Какова природа этих универсалий? Отражают ли они какие-то глубокие свойства эволюции или это просто статистические эффекты, не имеющие отношения к пониманию биологических явлений? Здесь и далее в этой книге (гл. 13) будет отстаиваться точка зрения на эти универсалии как на нетривиальные, характерные и биологически значимые тенденции, хотя они отражают только одну из двух (а возможно, и большего числа) дополнительных (в смысле принципа дополнительности Бора) составляющих эволюции жизни[44].
Во-первых, как это уже отмечалось и теперь совершенно очевидно, все эти универсалии зависимы от поведения совокупностей генов, фундаментальных единиц эволюции, рассматриваемых как статистические ансамбли. Таким образом, эти универсальные зависимости и распределения являются эмергентными свойствами биологических систем, то есть свойствами, проявляющимися в результате того, что эти системы состоят из многочисленных (достаточно многочисленных для проявления устойчивых статистических закономерностей) элементов (генов или белков, в зависимости от контекста), слабо взаимодействующих друг с другом (если сравнивать эти взаимодействия с теми, которые поддерживают целостность самих этих элементов).
Во-вторых, как мы уже видели, современный эволюционный анализ не останавливается на демонстрации существования универсальных понятий и законов. По крайней мере некоторые ключевые универсалии, такие как распределение эволюционных скоростей, отрицательные корреляции между скоростью эволюции и экспрессией и распределение численности паралогичных семейств, были теоретически выведены в рамках простых, но достаточно детализированных, формальных моделей эволюции. Способность простых моделей, в которых в качестве элементарных событий рассматриваются наиболее общие эволюционные процессы (такие как дупликация и утрата генов), объяснять геномные универсалии убеждает в том, что эти универсалии отражают существенные черты эволюции.
Третье, и, возможно, наиболее важное, замечание о новой парадигме понимания эволюции, которую мы пытаемся здесь обрисовать, состоит в том, что порождающие модели для общегеномных универсалий либо совсем не используют понятие отбора, либо используют только понятие очищающего (стабилизирующего) отбора. Эта форма отбора направлена на сохранение статуса-кво и наблюдается для укладки белковых молекул, для распределения численности генных семейств и для универсальной зависимости численности функциональных классов генов от общего числа генов (Koonin and Wolf, 2010b).
Аналогия между эволюционным процессом и статистической физикой не ограничена существованием универсальных зависимостей и распределений, некоторые из которых могут быть выведены в рамках простых моделей. Возможно также составить схему детального соответствия ключевых параметров этих двух областей (Barton and Coe, 2009; Sella and Hirsh, 2005). Такой параметр состояния (степень свободы), как положение частицы, в этой схеме является аналогом либо состояния сайта в нуклеотидной или белковой последовательности, либо состояния гена в геноме (в зависимости от уровня моделирования эволюции), и тогда параметрам скорости эволюции для сайта или гена будет соответствовать скорость частицы. Более того, значение эффективной численности популяции будет очевидно аналогичным значению температуры в статистической физике, а приспособленность будет соответствовать свободной энергии.
Во-первых, как это уже отмечалось и теперь совершенно очевидно, все эти универсалии зависимы от поведения совокупностей генов, фундаментальных единиц эволюции, рассматриваемых как статистические ансамбли. Таким образом, эти универсальные зависимости и распределения являются эмергентными свойствами биологических систем, то есть свойствами, проявляющимися в результате того, что эти системы состоят из многочисленных (достаточно многочисленных для проявления устойчивых статистических закономерностей) элементов (генов или белков, в зависимости от контекста), слабо взаимодействующих друг с другом (если сравнивать эти взаимодействия с теми, которые поддерживают целостность самих этих элементов).
Во-вторых, как мы уже видели, современный эволюционный анализ не останавливается на демонстрации существования универсальных понятий и законов. По крайней мере некоторые ключевые универсалии, такие как распределение эволюционных скоростей, отрицательные корреляции между скоростью эволюции и экспрессией и распределение численности паралогичных семейств, были теоретически выведены в рамках простых, но достаточно детализированных, формальных моделей эволюции. Способность простых моделей, в которых в качестве элементарных событий рассматриваются наиболее общие эволюционные процессы (такие как дупликация и утрата генов), объяснять геномные универсалии убеждает в том, что эти универсалии отражают существенные черты эволюции.
Третье, и, возможно, наиболее важное, замечание о новой парадигме понимания эволюции, которую мы пытаемся здесь обрисовать, состоит в том, что порождающие модели для общегеномных универсалий либо совсем не используют понятие отбора, либо используют только понятие очищающего (стабилизирующего) отбора. Эта форма отбора направлена на сохранение статуса-кво и наблюдается для укладки белковых молекул, для распределения численности генных семейств и для универсальной зависимости численности функциональных классов генов от общего числа генов (Koonin and Wolf, 2010b).
Аналогия между эволюционным процессом и статистической физикой не ограничена существованием универсальных зависимостей и распределений, некоторые из которых могут быть выведены в рамках простых моделей. Возможно также составить схему детального соответствия ключевых параметров этих двух областей (Barton and Coe, 2009; Sella and Hirsh, 2005). Такой параметр состояния (степень свободы), как положение частицы, в этой схеме является аналогом либо состояния сайта в нуклеотидной или белковой последовательности, либо состояния гена в геноме (в зависимости от уровня моделирования эволюции), и тогда параметрам скорости эволюции для сайта или гена будет соответствовать скорость частицы. Более того, значение эффективной численности популяции будет очевидно аналогичным значению температуры в статистической физике, а приспособленность будет соответствовать свободной энергии.
Краткий обзор и перспектива: о природе эволюционного процесса
Результаты взаимопроникновения сравнительной геномики и системной биологии, обсуждаемые в данной главе, приводят нас к следующему ключевому обобщению.
Многие, чтобы не сказать все, общие закономерности геномной и молекулярно-фенотипической эволюций описываются стохастическими процессами, основанными на принципе подверженной ошибкам репликации и ограниченными очищающим отбором, который поддерживает существующую общую (но не специфическую) архитектуру генома и устройства клеток.
Это обобщение не следует понимать как исключение адаптации из числа важнейших эволюционных понятий. Разумеется, адаптация – это общее и неотъемлемое явление в эволюции всех форм жизни. Тем не менее становится все более ясным, что общие количественные характеристики геномной архитектуры, функционирования и эволюции в первую очередь определяются неадаптивными, стохастическими процессами. Адаптация только модулирует эти закономерности. Здесь становится очень соблазнительным провести вполне очевидную параллель с нейтральной теорией Кимуры. В ходе высокоуровневого анализа геномных и молекулярно-фенотипических параметров мы начинаем различать контуры «неонейтрализма» (см. также гл. 8).
Аналогия между эволюцией и стохастическими физическими процессами ни в коем случае не отрицает метафоры «эволюции как мастерового» Жакоба. Напротив, новые открытия в эволюционной геномике прекрасно вписываются в это представление об эволюции: естественный отбор (адаптивный компонент эволюции) представляет собой процесс «латания», не полную перестройку или создание нового объекта, а добавление к существующему новых частей из уже имеющихся подручных материалов. Таким образом, первичная форма отбора – это очищающий отбор, который поддерживает статус-кво. Это обобщение имеет довольно удивительное, но неизбежное следствие: большая часть наиболее значимых событий во всей истории жизни произошла в течение первых нескольких сотен миллионов лет существования жизни на Земле, до появления современного типа клеток. Этот период в истории жизни должен был качественно отличаться от всей остальной эволюции; есть основания считать, что важнейшее достижение эволюции – это появление клетки, все остальное уже не так важно. Мы будем обсуждать происхождение жизни с этой точки зрения в главах 11 и 12 и вернемся к обсуждению общей природы эволюции в главе 13.
Параллели между эволюционной биологией и статистической физикой оказались точными и фундаментальными до такой степени, что кажется вполне справедливым заключение о том, что это не аналогии, а проявление общих статистических принципов (если не сказать законов) поведения больших ансамблей слабовзаимодействующих объектов[45]. Как в физике, так и в эволюционной биологии такие ансамбли (например, идеальный газ в физике и геном как сумма генов в биологии) являются идеализациями. В реальности отклонения от поведения, которое предсказывается простыми статистическими моделями, неизбежны и значимы. В эволюционной биологии такие отклонения, кроме всего прочего, вызываются различными взаимодействиями генов, что приводит к неожиданным эффектам, таким как отсутствие строгой корреляции между биологической значимостью гена и скоростью его эволюции. Тем не менее существенный эвристический потенциал прямого статистического подхода в объяснении по крайней мере некоторых фундаментальных свойств как физических, так и биологических процессов неоспорим.
Многие, чтобы не сказать все, общие закономерности геномной и молекулярно-фенотипической эволюций описываются стохастическими процессами, основанными на принципе подверженной ошибкам репликации и ограниченными очищающим отбором, который поддерживает существующую общую (но не специфическую) архитектуру генома и устройства клеток.
Это обобщение не следует понимать как исключение адаптации из числа важнейших эволюционных понятий. Разумеется, адаптация – это общее и неотъемлемое явление в эволюции всех форм жизни. Тем не менее становится все более ясным, что общие количественные характеристики геномной архитектуры, функционирования и эволюции в первую очередь определяются неадаптивными, стохастическими процессами. Адаптация только модулирует эти закономерности. Здесь становится очень соблазнительным провести вполне очевидную параллель с нейтральной теорией Кимуры. В ходе высокоуровневого анализа геномных и молекулярно-фенотипических параметров мы начинаем различать контуры «неонейтрализма» (см. также гл. 8).
Аналогия между эволюцией и стохастическими физическими процессами ни в коем случае не отрицает метафоры «эволюции как мастерового» Жакоба. Напротив, новые открытия в эволюционной геномике прекрасно вписываются в это представление об эволюции: естественный отбор (адаптивный компонент эволюции) представляет собой процесс «латания», не полную перестройку или создание нового объекта, а добавление к существующему новых частей из уже имеющихся подручных материалов. Таким образом, первичная форма отбора – это очищающий отбор, который поддерживает статус-кво. Это обобщение имеет довольно удивительное, но неизбежное следствие: большая часть наиболее значимых событий во всей истории жизни произошла в течение первых нескольких сотен миллионов лет существования жизни на Земле, до появления современного типа клеток. Этот период в истории жизни должен был качественно отличаться от всей остальной эволюции; есть основания считать, что важнейшее достижение эволюции – это появление клетки, все остальное уже не так важно. Мы будем обсуждать происхождение жизни с этой точки зрения в главах 11 и 12 и вернемся к обсуждению общей природы эволюции в главе 13.
Параллели между эволюционной биологией и статистической физикой оказались точными и фундаментальными до такой степени, что кажется вполне справедливым заключение о том, что это не аналогии, а проявление общих статистических принципов (если не сказать законов) поведения больших ансамблей слабовзаимодействующих объектов[45]. Как в физике, так и в эволюционной биологии такие ансамбли (например, идеальный газ в физике и геном как сумма генов в биологии) являются идеализациями. В реальности отклонения от поведения, которое предсказывается простыми статистическими моделями, неизбежны и значимы. В эволюционной биологии такие отклонения, кроме всего прочего, вызываются различными взаимодействиями генов, что приводит к неожиданным эффектам, таким как отсутствие строгой корреляции между биологической значимостью гена и скоростью его эволюции. Тем не менее существенный эвристический потенциал прямого статистического подхода в объяснении по крайней мере некоторых фундаментальных свойств как физических, так и биологических процессов неоспорим.
Рекомендуемая дополнительная литература[46]
Barabasi, A. L., and Z. N. Oltvai. (2004) Network Biology: Understanding the Cell’s Functional Organization. // Nature Reviews Genetics 5: 101–113.
Обзор свойств биологических сетей с акцентом на масштабной инвариантности.
Barton, N. H., and J. B. Coe. (2009) On the Application of Statistical Physics to Evolutionary Biology. Journal of Theoretical Biology 259: 317–324.
Технически сложная, но важная работа по термодинамическому подходу в эволюционной биологии.
Drummond, D. A., and C. O. Wilke. (2009) The Evolutionary Consequences of Erroneous Protein Synthesis. Nature Reviews Genetics 10: 715–724.
Критический обзор концепции эволюции, ограниченной ошибками трансляции и ошибками укладки белка.
Lobkovsky, A. E., Y. I. Wolf, and E. V. Koonin. (2010) Universal Distribution of Protein Evolution Rates As a Consequence of Protein Folding Physics. Proceedings of the National Academy of Sciences USA 107: 2,983—2,988.
В этой работе эволюционная динамика выводится в рамках простой модели укладки белка и с хорошей точностью воспроизводится универсальное распределение эволюционных скоростей.
Koonin, E. V., and Y. I. Wolf. (2006) Evolutionary Systems Biology: Links Between Gene Evolution and Function // Current Opinion in Biotechnology 17: 481–487.
Обзор корреляций между эволюционными и молекулярно-фенотипическими параметрами.
Koonin, E. V., Y. I. Wolf, and G. P. Karev. (2002) The Structure of the Protein Universe and Genome Evolution // Nature 420: 218–223.
Обсуждение универсальных распределений и зависимостей с акцентом на роли стохастических процессов и принципе предпочтительного присоединения.
Molina, N., and E. van Nimwegen. (2009) Scaling Laws in Functional Genome Content Across Prokaryotic Clades and Lifestyles // Trends in Genetics 25: 243–247.
Последние данные по универсальным степенным функциям для различных функциональных классов генов.
Sella, G., and A. E. Hirsh. (2005) The Application of Statistical Physics to Evolutionary Biology // Proceedings of the National Academy of Sciences USA 102: 9,541—9,546.
Богатая идеями статья, детально описывающая формальную аналогию между статистической физикой и эволюционной динамикой.
Schroedinger, Erwin. (1944/1992) What Is Life?: With «Mind and Matter» and «Autobiographical Sketches». Cambridge, MA: Cambridge University Press. [Шредингер Э. Что такое жизнь? Физический аспект живой клетки / Пер. с англ. 3-е изд. Ижевск: РХД, 2002.
Небольшая классическая книга, которую можно порекомендовать еще раз как взгляд на биологию с физической точки зрения, не так уж и изменившийся со времен Шредингера.
Обзор свойств биологических сетей с акцентом на масштабной инвариантности.
Barton, N. H., and J. B. Coe. (2009) On the Application of Statistical Physics to Evolutionary Biology. Journal of Theoretical Biology 259: 317–324.
Технически сложная, но важная работа по термодинамическому подходу в эволюционной биологии.
Drummond, D. A., and C. O. Wilke. (2009) The Evolutionary Consequences of Erroneous Protein Synthesis. Nature Reviews Genetics 10: 715–724.
Критический обзор концепции эволюции, ограниченной ошибками трансляции и ошибками укладки белка.
Lobkovsky, A. E., Y. I. Wolf, and E. V. Koonin. (2010) Universal Distribution of Protein Evolution Rates As a Consequence of Protein Folding Physics. Proceedings of the National Academy of Sciences USA 107: 2,983—2,988.
В этой работе эволюционная динамика выводится в рамках простой модели укладки белка и с хорошей точностью воспроизводится универсальное распределение эволюционных скоростей.
Koonin, E. V., and Y. I. Wolf. (2006) Evolutionary Systems Biology: Links Between Gene Evolution and Function // Current Opinion in Biotechnology 17: 481–487.
Обзор корреляций между эволюционными и молекулярно-фенотипическими параметрами.
Koonin, E. V., Y. I. Wolf, and G. P. Karev. (2002) The Structure of the Protein Universe and Genome Evolution // Nature 420: 218–223.
Обсуждение универсальных распределений и зависимостей с акцентом на роли стохастических процессов и принципе предпочтительного присоединения.
Molina, N., and E. van Nimwegen. (2009) Scaling Laws in Functional Genome Content Across Prokaryotic Clades and Lifestyles // Trends in Genetics 25: 243–247.
Последние данные по универсальным степенным функциям для различных функциональных классов генов.
Sella, G., and A. E. Hirsh. (2005) The Application of Statistical Physics to Evolutionary Biology // Proceedings of the National Academy of Sciences USA 102: 9,541—9,546.
Богатая идеями статья, детально описывающая формальную аналогию между статистической физикой и эволюционной динамикой.
Schroedinger, Erwin. (1944/1992) What Is Life?: With «Mind and Matter» and «Autobiographical Sketches». Cambridge, MA: Cambridge University Press. [Шредингер Э. Что такое жизнь? Физический аспект живой клетки / Пер. с англ. 3-е изд. Ижевск: РХД, 2002.
Небольшая классическая книга, которую можно порекомендовать еще раз как взгляд на биологию с физической точки зрения, не так уж и изменившийся со времен Шредингера.
Глава 5. Сетевая геномика мира прокариот: вертикальные и горизонтальные потоки генов, мобиломы и динамика пангеномов
Когда Дарвин писал об эволюции, он имел в виду животных и растения, по крайней мере он использовал эти сложные многоклеточные организмы во всех своих конкретных примерах. Одноклеточные формы жизни практически не упоминаются в «Происхождении видов…» или любой другой книге Дарвина. В любом случае, учитывая, что Дарвин серьезно обсуждал происхождение всех существующих ныне видов от одной или нескольких предковых форм (см. гл. 2 и 11), он должен был исходить из того, что эти предки были одноклеточными[47]. Эрнст Геккель, плодовитый немецкий последователь Дарвина, поместил протист (одноклеточных эукариотов, часто называемых этим термином даже сейчас) и дробянок (ныне известных как прокариоты – бактерии и археи) в основании своего монументального древа жизни, первого из подобных деревьев, которое было населено реальными жизненными формами. Естественно, животные доминировали на дереве Геккеля, в то время как протисты и дробянки располагались на неопределенных позициях поблизости от корня.
Вездесущесть и важность бактерий в биосфере постепенно становились очевидными параллельно с развитием эволюционной биологии, вначале благодаря полным драматизма исследованиям бактериальных патогенов, а позднее в результате достижений экологической микробиологии. Достаточно рано микробиологи показали, что бактерии в буквальном смысле являются основным действующим началом в биосфере. Подавляющее большинство живых клеток на нашей планете – это именно бактерии, они демонстрируют наибольшее биохимическое разнообразие среди всех организмов и являются главной геохимической силой. Однако, несмотря на биологическую важность и поразительное биохимическое и экологическое разнообразие микробов и огромный прогресс микробиологии в середине XX столетия (в качестве примеров можно упомянуть открытие антибиотиков и демонстрацию химической природы генетического материала бактерий), микробиология ничего не внесла в СТЭ и не была эволюционной дисциплиной на протяжении большей части этого столетия. Не то чтобы микробиологи совсем не думали об эволюции, но все их попытки расшифровать эволюционные взаимоотношения между бактериями, используя морфологию клеток, а также метаболические и фенотипические характеристики, и на основе этих признаков построить филогенетическую таксономию приводили к несовместимым и неприемлемым результатам. Весьма любопытно, что приблизительно в то время, когда происходила консолидация СТЭ, ведущие микробиологии того времени, включая Роджера Стейнира и Корнелиуса Ван Нейла, пришли к выводу, что, если какие-то эволюционные процессы и происходят в микромире, описать их и как-либо применить в сфере таксономии микробов и микробиологии вообще практически невозможно (Stanier and Van Niel, 1962; Van Niel, 1955).
Как отмечено в главе 3, все резко изменилось в 1977 году, когда Карл Вёзе с сотрудниками применил филогенетический анализ рРНК как основной метод изучения эволюции микробов и создания их таксономии (Woese, 1987). Возможности новой методологии были эффектно продемонстрированы открытием архей, по-видимому, первым крупным открытием в биологии, которое было сделано исключительно на основе анализа нуклеотидных последовательностей. За этим прорывом последовал период «бури и натиска» 1980-х и начала 1990-х годов, когда филогения рРНК была успешно применена для прояснения взаимоотношений среди многих групп прокариот. Среди молекулярных эволюционистов того времени превалировало мнение, что в принципе эти методы позволят точно реконструировать эволюцию микробов.
Однако дивный новый мир микробной эволюции оказался недолговечным – эволюционная геномика вновь запутала картину самым неожиданным образом. Первый полный бактериальный геном был секвенирован в 1995 году, а первый геном археи – в 1996-м[48]. Вскоре после этого прорыва установился экспоненциальный темп секвенирования геномов со временем удвоения около 20 месяцев для бактерий и около 34 месяцев для архей (см. рис. 3–1). Сравнительный анализ сотен секвенированных бактериальных геномов и десятков геномов архей привел к важнейшему выводу: микробы определенно эволюционируют, но их эволюция сильно отличается от той, что описана СТЭ (Doolittle, 1999b; Woese and Goldenfeld, 2009). Ключевым стало осознание того, что геномы прокариот ведут себя не так, как если бы они были стабильными, точно наследуемыми носителями генетической информации организма (вида). Геномы микробов оказались чрезвычайно динамичными, неоднородными образованиями, которые относительно стабильны лишь на коротких интервалах времени, имеют свою характерную скорость распада и существуют в динамическом равновесии между различными формами жизни, которые отличаются по принципам геномной организации. В «мире прокариот» эти взаимосвязанные и постоянно взаимодействующие формы жизни включают не только бактерии и археи, но также различные плазмиды, вирусы и другие мобильные элементы. В этой новой, динамической парадигме прокариотической эволюции традиционная концепция видов с четко определенным, стабильным геномом теряет существенную, если не большую часть своей применимости (Doolittle and Zhaxybayeva, 2009). Становится осмысленнее говорить о сериях «пангеномов» на всех уровнях, от пангенома, например, Escherichia coli или любого другого «вида» бактерий или архей, до пангенома всех прокариот (Lapierre and Gogarten, 2009; Mira et al., 2010).
В главе 3 мы уже обсуждали важные аспекты структуры генетической вселенной прокариот. Она рассматривалась в основном как сложный статичный объект, то есть в терминах распределения различных существенных переменных. В этой главе мы также рассматриваем распределения, но в основном пытаемся встать на динамическую точку зрения и исследовать мир прокариот в терминах потоков генов и взаимодействия между репликонами.
Вездесущесть и важность бактерий в биосфере постепенно становились очевидными параллельно с развитием эволюционной биологии, вначале благодаря полным драматизма исследованиям бактериальных патогенов, а позднее в результате достижений экологической микробиологии. Достаточно рано микробиологи показали, что бактерии в буквальном смысле являются основным действующим началом в биосфере. Подавляющее большинство живых клеток на нашей планете – это именно бактерии, они демонстрируют наибольшее биохимическое разнообразие среди всех организмов и являются главной геохимической силой. Однако, несмотря на биологическую важность и поразительное биохимическое и экологическое разнообразие микробов и огромный прогресс микробиологии в середине XX столетия (в качестве примеров можно упомянуть открытие антибиотиков и демонстрацию химической природы генетического материала бактерий), микробиология ничего не внесла в СТЭ и не была эволюционной дисциплиной на протяжении большей части этого столетия. Не то чтобы микробиологи совсем не думали об эволюции, но все их попытки расшифровать эволюционные взаимоотношения между бактериями, используя морфологию клеток, а также метаболические и фенотипические характеристики, и на основе этих признаков построить филогенетическую таксономию приводили к несовместимым и неприемлемым результатам. Весьма любопытно, что приблизительно в то время, когда происходила консолидация СТЭ, ведущие микробиологии того времени, включая Роджера Стейнира и Корнелиуса Ван Нейла, пришли к выводу, что, если какие-то эволюционные процессы и происходят в микромире, описать их и как-либо применить в сфере таксономии микробов и микробиологии вообще практически невозможно (Stanier and Van Niel, 1962; Van Niel, 1955).
Как отмечено в главе 3, все резко изменилось в 1977 году, когда Карл Вёзе с сотрудниками применил филогенетический анализ рРНК как основной метод изучения эволюции микробов и создания их таксономии (Woese, 1987). Возможности новой методологии были эффектно продемонстрированы открытием архей, по-видимому, первым крупным открытием в биологии, которое было сделано исключительно на основе анализа нуклеотидных последовательностей. За этим прорывом последовал период «бури и натиска» 1980-х и начала 1990-х годов, когда филогения рРНК была успешно применена для прояснения взаимоотношений среди многих групп прокариот. Среди молекулярных эволюционистов того времени превалировало мнение, что в принципе эти методы позволят точно реконструировать эволюцию микробов.
Однако дивный новый мир микробной эволюции оказался недолговечным – эволюционная геномика вновь запутала картину самым неожиданным образом. Первый полный бактериальный геном был секвенирован в 1995 году, а первый геном археи – в 1996-м[48]. Вскоре после этого прорыва установился экспоненциальный темп секвенирования геномов со временем удвоения около 20 месяцев для бактерий и около 34 месяцев для архей (см. рис. 3–1). Сравнительный анализ сотен секвенированных бактериальных геномов и десятков геномов архей привел к важнейшему выводу: микробы определенно эволюционируют, но их эволюция сильно отличается от той, что описана СТЭ (Doolittle, 1999b; Woese and Goldenfeld, 2009). Ключевым стало осознание того, что геномы прокариот ведут себя не так, как если бы они были стабильными, точно наследуемыми носителями генетической информации организма (вида). Геномы микробов оказались чрезвычайно динамичными, неоднородными образованиями, которые относительно стабильны лишь на коротких интервалах времени, имеют свою характерную скорость распада и существуют в динамическом равновесии между различными формами жизни, которые отличаются по принципам геномной организации. В «мире прокариот» эти взаимосвязанные и постоянно взаимодействующие формы жизни включают не только бактерии и археи, но также различные плазмиды, вирусы и другие мобильные элементы. В этой новой, динамической парадигме прокариотической эволюции традиционная концепция видов с четко определенным, стабильным геномом теряет существенную, если не большую часть своей применимости (Doolittle and Zhaxybayeva, 2009). Становится осмысленнее говорить о сериях «пангеномов» на всех уровнях, от пангенома, например, Escherichia coli или любого другого «вида» бактерий или архей, до пангенома всех прокариот (Lapierre and Gogarten, 2009; Mira et al., 2010).
В главе 3 мы уже обсуждали важные аспекты структуры генетической вселенной прокариот. Она рассматривалась в основном как сложный статичный объект, то есть в терминах распределения различных существенных переменных. В этой главе мы также рассматриваем распределения, но в основном пытаемся встать на динамическую точку зрения и исследовать мир прокариот в терминах потоков генов и взаимодействия между репликонами.
Размер и общая организация бактериальных и архейных геномов
Несмотря на огромные различия в образе жизни, а также метаболической и геномной организации, бактериальные и архейные геномы демонстрируют легко различимые общие архитектурные принципы (см. обзор в гл. 3). Секвенированные бактериальные и архейные геномы охватывают два порядка величины по размерам от около 144 Кб для внутриклеточного симбионта Hodgkinia cicadicola до примерно 13 Мб для обитающей в почве бактерии Sorangium cellulosum (Koonin and Wolf, 2008b). Примечательно, что бактерии демонстрируют бимодальное распределение размеров генома[49] с пиком в районе примерно 5 Мб и дополнительным плато в районе примерно 2 Мб (см. рис. 5–1). Хотя существует много геномов промежуточного размера, это распределение предполагает существование двух в достаточной степени разделенных классов бактерий с «малым» и «большим» геномами. К этим наблюдениям нужно относиться с известной осторожностью, так как они могут быть артефактом, обусловленным предпочтительным секвенированием небольших геномов (в первую очередь бактериальных патогенов), но с ростом числа секвенированных геномов такое объяснение становится все менее удовлетворительным.
Археи демонстрируют более узкое, но также сложное распределение размеров генома от примерно 0,5 Мб у паразита/симбионта Nanoarchaeum equitans до примерно 5,5 Мб у Methanosarcina barkeri, с острым пиком в районе 2 Мб, который практически точно соответствует расположению плато бактериальных геномов малого размера, вторым небольшим пиком около 3 Мб и тяжелым хвостом, соответствующим геномам большего размера (см. рис. 5–1). При этом смещения в базе данных опять могут быть существенными, так как в настоящее время геномов архей секвенировано примерно на порядок меньше, чем геномов бактерий, так что пока может быть еще просто недостаточно данных для выявления истинной формы распределения размеров геномов. Однако более вероятно, что археи действительно являются менее разнородной группой, как будет обсуждаться далее в данном разделе.
Все очень маленькие (менее 1 Мб) геномы бактерий и архей принадлежат бактериям-паразитам и внутриклеточным симбионтам эукариот и единственной известной архее-паразиту (или симбионту) Nanoarchaeum equitans, которая живет за счет другой археи, Ignicoccus hospitalis. Таким образом, кажется все более вероятным, что минимальный размер генома свободно живущего прокариота, по крайней мере автотрофа, который не зависит от других форм жизни для добывания пищи, немного превышает 1 Мб. Текущий рекорд редукции генома среди свободно живущих клеток, около 1,3 Мб, принадлежит фотосинтезирующей морской альфа-протеобактерии Pelagibacter ubique (SAR11), которая также является наиболее распространенной из известных клеточных форм жизни на Земле (Giovannoni et al., 2005). (Связь между размером популяции и размером генома потенциально важна, мы вернемся к этому вопросу в гл. 8.)
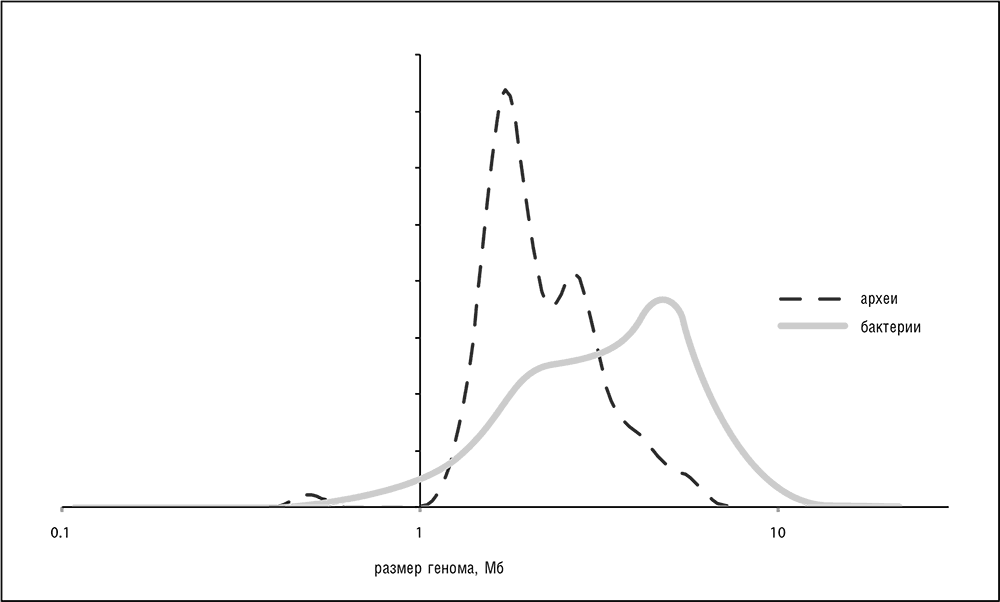 Рис. 5–1. Распределение размеров геномов среди бактерий и архей.
Рис. 5–1. Распределение размеров геномов среди бактерий и архей.
Как мы уже обсуждали в главе 3, бактериальные и архейные геномы характеризуются высокой плотностью белок-кодирующих генов, которые занимают большую часть ДНК. Бактериальные и архейные геномы демонстрируют одномодальное и довольно острое распределение плотности генов, большей частью 0,8–1,2 гена на Кб геномной ДНК (отсюда предельно простое эмпирическое правило: 1 ген на 1000 пар нуклеотидов). Архейное распределение по сравнению с бактериальным сдвинуто в сторону более высоких плотностей, таким образом, в среднем архейные геномы даже более компактны, чем бактериальные. Похоже, что как кодирующие, так и межгенные области у архей немного короче по сравнению с бактериями.
Таким образом, археи и бактерии весьма похожи в смысле характерных размеров и общей архитектуры геномов, но резко отличаются от эукариот, которые охватывают много больший интервал размеров генома, имеют белок-кодирующие гены, часто прерываемые интронами, и более длинные межгенные промежутки (см. гл. 8). Эти общие признаки бактерий и архей подтверждают концепцию «прокариотного принципа организации генома» (см. более подробно ниже).
Археи демонстрируют более узкое, но также сложное распределение размеров генома от примерно 0,5 Мб у паразита/симбионта Nanoarchaeum equitans до примерно 5,5 Мб у Methanosarcina barkeri, с острым пиком в районе 2 Мб, который практически точно соответствует расположению плато бактериальных геномов малого размера, вторым небольшим пиком около 3 Мб и тяжелым хвостом, соответствующим геномам большего размера (см. рис. 5–1). При этом смещения в базе данных опять могут быть существенными, так как в настоящее время геномов архей секвенировано примерно на порядок меньше, чем геномов бактерий, так что пока может быть еще просто недостаточно данных для выявления истинной формы распределения размеров геномов. Однако более вероятно, что археи действительно являются менее разнородной группой, как будет обсуждаться далее в данном разделе.
Все очень маленькие (менее 1 Мб) геномы бактерий и архей принадлежат бактериям-паразитам и внутриклеточным симбионтам эукариот и единственной известной архее-паразиту (или симбионту) Nanoarchaeum equitans, которая живет за счет другой археи, Ignicoccus hospitalis. Таким образом, кажется все более вероятным, что минимальный размер генома свободно живущего прокариота, по крайней мере автотрофа, который не зависит от других форм жизни для добывания пищи, немного превышает 1 Мб. Текущий рекорд редукции генома среди свободно живущих клеток, около 1,3 Мб, принадлежит фотосинтезирующей морской альфа-протеобактерии Pelagibacter ubique (SAR11), которая также является наиболее распространенной из известных клеточных форм жизни на Земле (Giovannoni et al., 2005). (Связь между размером популяции и размером генома потенциально важна, мы вернемся к этому вопросу в гл. 8.)
Как мы уже обсуждали в главе 3, бактериальные и архейные геномы характеризуются высокой плотностью белок-кодирующих генов, которые занимают большую часть ДНК. Бактериальные и архейные геномы демонстрируют одномодальное и довольно острое распределение плотности генов, большей частью 0,8–1,2 гена на Кб геномной ДНК (отсюда предельно простое эмпирическое правило: 1 ген на 1000 пар нуклеотидов). Архейное распределение по сравнению с бактериальным сдвинуто в сторону более высоких плотностей, таким образом, в среднем архейные геномы даже более компактны, чем бактериальные. Похоже, что как кодирующие, так и межгенные области у архей немного короче по сравнению с бактериями.
Таким образом, археи и бактерии весьма похожи в смысле характерных размеров и общей архитектуры геномов, но резко отличаются от эукариот, которые охватывают много больший интервал размеров генома, имеют белок-кодирующие гены, часто прерываемые интронами, и более длинные межгенные промежутки (см. гл. 8). Эти общие признаки бактерий и архей подтверждают концепцию «прокариотного принципа организации генома» (см. более подробно ниже).