• Дарвин предложил конкретный, прямой и доходчивый механизм эволюции, представляющий собой взаимодействие между наследственной изменчивостью и естественным отбором, в целом описываемое как выживание наиболее приспособленных.
• Дарвин смело расширил идеи эволюции на всю историю земной жизни, которая, как он полагал, может быть представлена величественным древом (знаменитая единственная иллюстрация в «Происхождении…»), и даже утверждал, что все существующие формы жизни происходят от единого общего предка.
Общая и обладающая огромной предсказательной силой модель эволюции, предложенная Дарвином, явилась резким контрастом к эволюционным идеям его предшественников, особенно Ламарка и Лайеля, которые рассматривали преимущественно или даже исключительно внутривидовые эволюционные изменения. Четвертое значительное достижение Дарвина связано не столько с научным содержанием его работы, сколько с формой ее изложения. Главным образом в связи с вполне понятной срочностью, вызванной соперничеством с Уоллесом, Дарвин представил свой труд в виде небольшой и легко читаемой даже для неспециалиста книги, которая, несмотря на это, содержала скрупулезно и тщательно собранные доводы. Благодаря этим принципиальным достижениям, Дарвин не просто опубликовал очередную книгу об эволюции, но полностью изменил лицо науки. Сразу же после публикации «Происхождения…» большинство биологов и даже просто образованная часть общества признали эту работу как заслуживающее доверие естественно-научное объяснение возникновения многообразия форм жизни, и это послужило динамичной основой для дальнейших теоретических построений[4].
Рассматривая труд Дарвина с более отвлеченной позиции, которая является основной в этой книге, необходимо особо отметить, что Дарвин, похоже, первым обнаружил определяющее взаимодействие между случаем и направленностью (неизбежностью) в эволюции. В соответствии с идеей Дарвина, изменчивость почти полностью случайна, в то время как отбор является направленным и создает сложность. В этом Дарвин полностью противоположен Ламарку, который, в сущности, изгнал случайность из своей картины мира. В данной книге мы будем периодически возвращаться к этому ключевому конфликту мировоззрений.
Конечно, надо отдать должное предшественникам Дарвина – геологам и эволюционным биологам, однако Дарвин, несомненно, был первым ученым, который включил возможность эволюционных изменений (и, косвенно, происхождение) всей Вселенной в сферу явлений природы, подлежащих рациональному изучению. Другими словами, Дарвин положил начало научному изучению стрелы времени – то есть асимметричных во времени, необратимых процессов. Таким образом, он подготовил почву не только для развития биологии, но также для создания современной физики. Я полагаю, что знаменитый физик Людвиг Больцман, основатель статистической термодинамики и автор современной концепции энтропии, имел все основания назвать Дарвина «великим физиком», что может показаться парадоксальным, учитывая, что Дарвин крайне мало знал физику и математику. Пожалуй, и наш современник, философ Дэниел Деннет не так уж преувеличил, утверждая, что дарвиновская идея естественного отбора – величайшая идея в истории человечества (Dennett, 1996).
Конечно, эволюционное учение Дарвина со времени публикации «Происхождения…» и по меньшей мере до конца XIX века сталкивалось с острыми проблемами, всерьез беспокоившими Дарвина и на тот момент казавшимися непреодолимыми большинству ученых. Во-первых, значительную трудность представляло собой определение возраста Земли, который во времена Дарвина был существенно занижен. Даже не принимая во внимание религиозные мифы о сотворении мира, наиболее точно возраст Земли, по мнению физиков XIX века (в частности, лорда Кельвина), оценивался в 100 миллионов лет. Такого промежутка времени было явно недостаточно для эволюции жизни в том виде, в каком ее представил Дарвин, то есть путем постепенного накопления небольших изменений. В целом, действительно, 100 миллионов лет очень мало для эволюции жизни в ее нынешнем многообразии, хотя никто в XIX веке не мог количественно оценить скорость дарвиновской эволюции. Эта проблема разрешилась спустя 20 лет после смерти Дарвина. После открытия радиоактивности в начале XX века ученые подсчитали, что охлаждение Земли от первичного раскаленного состояния заняло миллиарды лет, то есть примерно столько, сколько, по предположению Дарвина, требовалось для эволюции с помощью естественного отбора.
Во-вторых, еще больше вопросов вызывали механизмы наследственности и так называемый кошмар Дженкина. Так как во времена Дарвина еще не существовало теории дискретных наследственных детерминант (кроме малоизвестных статей Менделя), то было неясно, каким образом полезное благоприобретение может сохраниться в поколениях и закрепиться в эволюционирующей популяции, не растворяясь и не теряясь. Очевидно, сам Дарвин не обратил внимания на эту проблему в своей теории, когда писал «Происхождение…», однако о ней сообщил Дарвину необычайно критичный читатель его работы, инженер Дженкин. Оглядываясь назад, трудно понять, почему Дарвин (или Дженкин, или Гексли) не принял в расчет решение, предлагаемое Менделем. Вместо этого Дарвин выдвинул куда более странное объяснение, так называемую теорию пангенеза, которую даже он сам, по-видимому, не принимал всерьез. Противоречие было устранено с рождением (или, вернее, повторным рождением) генетики, хотя поначалу ее значение для дарвинизма[5] было неочевидным (см. следующую главу).
Третья проблема, которую Дарвин полностью осознавал и блестяще исследовал, – это эволюция сложных структур (органов, по терминологии Дарвина), для работы которых необходимо соединение множества частей. Такие сложные органы представляли собой классическую головоломку для эволюционной биологии, которая в XX веке была выразительно названа неупрощаемой сложностью[6]. Конечно, сразу непонятно, как может происходить эволюция таких органов путем естественного отбора, если считать, что отдельные части органа или «частично укомплектованный» орган не функциональны. Дарвин решительно обратился к этой проблеме в одном из самых известных отрывков «Происхождения…», сценарии эволюции глаза. Он предложил логически безупречное, убедительное и неординарное решение: Дарвин предположил, что эволюция сложных органов идет через серию промежуточных стадий, каждая из которых частично выполняет функцию развивающегося сложного органа. Таким образом, эволюция глаза, по Дарвину, начинается с простого светочувствительного участка, через примитивные постепенно усложняющиеся структуры, подобные глазу, к полноценным, функциональным сложным глазам членистоногих и позвоночных. Необходимо отметить, что примитивные светочувствительные структуры, похожие на те, существование которых предположил Дарвин исходя из общих предположений, были впоследствии обнаружены, что по крайней мере частично подтверждает его сценарий и показывает, что в этом случае неупрощаемость сложной структуры иллюзорна. Однако, несмотря на убедительность схемы, предложенной Дарвином, к ней следует относиться трезво, как к частично подтверждаемому, но все же гипотетическому сценарию эволюции одного конкретного органа. Предположение Дарвина показало одну из возможных траекторий эволюции сложной структуры, но не решило главную проблему в целом. Эволюция сложных структур на разных уровнях является центральным вопросом биологии, поэтому мы будем возвращаться к нему много раз в этой книге.
Четвертый сложный вопрос дарвинизма является и самым глубоким. Эта главная проблема имеет непосредственное отношение к названию книги Дарвина и к подразумеваемой основной ее теме, то есть к происхождению видов, и, в общем смысле, к крупным эволюционным событиям, которые в настоящее время носят собирательное название макроэволюция. В значительном отрыве от названия книги те неоспоримые примеры эволюции, которые представил Дарвин, относились к возникновению новых внутривидовых различий, а не новых видов, не говоря уже о новых таксонах более высокого уровня. Эта проблема сохранялась долго после смерти Дарвина и существует даже сейчас, хотя частично она была решена сначала прогрессом палеонтологии, затем развитием теории видообразования при поддержке биогеографических данных, а затем, наиболее убедительно, сравнительной геномикой (см. гл. 2 и 3). К чести Дарвина и в отличие от критиков эволюции по сей день, он твердо стоял на своем перед лицом всех трудностей, благодаря своей непоколебимой вере в то, что, несмотря на возможные пробелы в его теории, ей нет никакой разумной альтернативы. Единственным слабым местом Дарвина оказалось включение неправдоподобной модели пангенеза в последующие издания «Происхождения…» как заплатки для маскировки кошмара Дженкина.
Генетика и «черный день» дарвинизма
Популяционная генетика, теорема Фишера, адаптивные ландшафты, генетический дрейф и «эволюционная тяга»
Положительный и очищающий (отрицательный) отбор: классификация форм отбора
• Дарвин смело расширил идеи эволюции на всю историю земной жизни, которая, как он полагал, может быть представлена величественным древом (знаменитая единственная иллюстрация в «Происхождении…»), и даже утверждал, что все существующие формы жизни происходят от единого общего предка.
Общая и обладающая огромной предсказательной силой модель эволюции, предложенная Дарвином, явилась резким контрастом к эволюционным идеям его предшественников, особенно Ламарка и Лайеля, которые рассматривали преимущественно или даже исключительно внутривидовые эволюционные изменения. Четвертое значительное достижение Дарвина связано не столько с научным содержанием его работы, сколько с формой ее изложения. Главным образом в связи с вполне понятной срочностью, вызванной соперничеством с Уоллесом, Дарвин представил свой труд в виде небольшой и легко читаемой даже для неспециалиста книги, которая, несмотря на это, содержала скрупулезно и тщательно собранные доводы. Благодаря этим принципиальным достижениям, Дарвин не просто опубликовал очередную книгу об эволюции, но полностью изменил лицо науки. Сразу же после публикации «Происхождения…» большинство биологов и даже просто образованная часть общества признали эту работу как заслуживающее доверие естественно-научное объяснение возникновения многообразия форм жизни, и это послужило динамичной основой для дальнейших теоретических построений[4].
Рассматривая труд Дарвина с более отвлеченной позиции, которая является основной в этой книге, необходимо особо отметить, что Дарвин, похоже, первым обнаружил определяющее взаимодействие между случаем и направленностью (неизбежностью) в эволюции. В соответствии с идеей Дарвина, изменчивость почти полностью случайна, в то время как отбор является направленным и создает сложность. В этом Дарвин полностью противоположен Ламарку, который, в сущности, изгнал случайность из своей картины мира. В данной книге мы будем периодически возвращаться к этому ключевому конфликту мировоззрений.
Конечно, надо отдать должное предшественникам Дарвина – геологам и эволюционным биологам, однако Дарвин, несомненно, был первым ученым, который включил возможность эволюционных изменений (и, косвенно, происхождение) всей Вселенной в сферу явлений природы, подлежащих рациональному изучению. Другими словами, Дарвин положил начало научному изучению стрелы времени – то есть асимметричных во времени, необратимых процессов. Таким образом, он подготовил почву не только для развития биологии, но также для создания современной физики. Я полагаю, что знаменитый физик Людвиг Больцман, основатель статистической термодинамики и автор современной концепции энтропии, имел все основания назвать Дарвина «великим физиком», что может показаться парадоксальным, учитывая, что Дарвин крайне мало знал физику и математику. Пожалуй, и наш современник, философ Дэниел Деннет не так уж преувеличил, утверждая, что дарвиновская идея естественного отбора – величайшая идея в истории человечества (Dennett, 1996).
Конечно, эволюционное учение Дарвина со времени публикации «Происхождения…» и по меньшей мере до конца XIX века сталкивалось с острыми проблемами, всерьез беспокоившими Дарвина и на тот момент казавшимися непреодолимыми большинству ученых. Во-первых, значительную трудность представляло собой определение возраста Земли, который во времена Дарвина был существенно занижен. Даже не принимая во внимание религиозные мифы о сотворении мира, наиболее точно возраст Земли, по мнению физиков XIX века (в частности, лорда Кельвина), оценивался в 100 миллионов лет. Такого промежутка времени было явно недостаточно для эволюции жизни в том виде, в каком ее представил Дарвин, то есть путем постепенного накопления небольших изменений. В целом, действительно, 100 миллионов лет очень мало для эволюции жизни в ее нынешнем многообразии, хотя никто в XIX веке не мог количественно оценить скорость дарвиновской эволюции. Эта проблема разрешилась спустя 20 лет после смерти Дарвина. После открытия радиоактивности в начале XX века ученые подсчитали, что охлаждение Земли от первичного раскаленного состояния заняло миллиарды лет, то есть примерно столько, сколько, по предположению Дарвина, требовалось для эволюции с помощью естественного отбора.
Во-вторых, еще больше вопросов вызывали механизмы наследственности и так называемый кошмар Дженкина. Так как во времена Дарвина еще не существовало теории дискретных наследственных детерминант (кроме малоизвестных статей Менделя), то было неясно, каким образом полезное благоприобретение может сохраниться в поколениях и закрепиться в эволюционирующей популяции, не растворяясь и не теряясь. Очевидно, сам Дарвин не обратил внимания на эту проблему в своей теории, когда писал «Происхождение…», однако о ней сообщил Дарвину необычайно критичный читатель его работы, инженер Дженкин. Оглядываясь назад, трудно понять, почему Дарвин (или Дженкин, или Гексли) не принял в расчет решение, предлагаемое Менделем. Вместо этого Дарвин выдвинул куда более странное объяснение, так называемую теорию пангенеза, которую даже он сам, по-видимому, не принимал всерьез. Противоречие было устранено с рождением (или, вернее, повторным рождением) генетики, хотя поначалу ее значение для дарвинизма[5] было неочевидным (см. следующую главу).
Третья проблема, которую Дарвин полностью осознавал и блестяще исследовал, – это эволюция сложных структур (органов, по терминологии Дарвина), для работы которых необходимо соединение множества частей. Такие сложные органы представляли собой классическую головоломку для эволюционной биологии, которая в XX веке была выразительно названа неупрощаемой сложностью[6]. Конечно, сразу непонятно, как может происходить эволюция таких органов путем естественного отбора, если считать, что отдельные части органа или «частично укомплектованный» орган не функциональны. Дарвин решительно обратился к этой проблеме в одном из самых известных отрывков «Происхождения…», сценарии эволюции глаза. Он предложил логически безупречное, убедительное и неординарное решение: Дарвин предположил, что эволюция сложных органов идет через серию промежуточных стадий, каждая из которых частично выполняет функцию развивающегося сложного органа. Таким образом, эволюция глаза, по Дарвину, начинается с простого светочувствительного участка, через примитивные постепенно усложняющиеся структуры, подобные глазу, к полноценным, функциональным сложным глазам членистоногих и позвоночных. Необходимо отметить, что примитивные светочувствительные структуры, похожие на те, существование которых предположил Дарвин исходя из общих предположений, были впоследствии обнаружены, что по крайней мере частично подтверждает его сценарий и показывает, что в этом случае неупрощаемость сложной структуры иллюзорна. Однако, несмотря на убедительность схемы, предложенной Дарвином, к ней следует относиться трезво, как к частично подтверждаемому, но все же гипотетическому сценарию эволюции одного конкретного органа. Предположение Дарвина показало одну из возможных траекторий эволюции сложной структуры, но не решило главную проблему в целом. Эволюция сложных структур на разных уровнях является центральным вопросом биологии, поэтому мы будем возвращаться к нему много раз в этой книге.
Четвертый сложный вопрос дарвинизма является и самым глубоким. Эта главная проблема имеет непосредственное отношение к названию книги Дарвина и к подразумеваемой основной ее теме, то есть к происхождению видов, и, в общем смысле, к крупным эволюционным событиям, которые в настоящее время носят собирательное название макроэволюция. В значительном отрыве от названия книги те неоспоримые примеры эволюции, которые представил Дарвин, относились к возникновению новых внутривидовых различий, а не новых видов, не говоря уже о новых таксонах более высокого уровня. Эта проблема сохранялась долго после смерти Дарвина и существует даже сейчас, хотя частично она была решена сначала прогрессом палеонтологии, затем развитием теории видообразования при поддержке биогеографических данных, а затем, наиболее убедительно, сравнительной геномикой (см. гл. 2 и 3). К чести Дарвина и в отличие от критиков эволюции по сей день, он твердо стоял на своем перед лицом всех трудностей, благодаря своей непоколебимой вере в то, что, несмотря на возможные пробелы в его теории, ей нет никакой разумной альтернативы. Единственным слабым местом Дарвина оказалось включение неправдоподобной модели пангенеза в последующие издания «Происхождения…» как заплатки для маскировки кошмара Дженкина.
Генетика и «черный день» дарвинизма
Существует легенда, что Дарвин прочитал работу Менделя, но не нашел ее интересной (возможно, из-за ограниченного знания немецкого языка). Сложно предположить, насколько изменилась бы история биологии, если бы Дарвин использовал идеи Менделя, которые теперь нам кажутся предельно простыми. Однако этого не произошло.
Еще удивительнее, что сам Мендель, очевидно хорошо знакомый с «Происхождением…»[7], не рассматривал свое открытие в контексте теории Дарвина. Ожидать установления этой жизненно важной связи пришлось не только до возрождения генетики на заре XX века, но также до появления популяционной генетики в 1920-х годах. Повторное открытие механизма наследования и рождение генетики дало мощный толчок развитию дарвинизма, так как выявление дискретных носителей наследственности устраняло кошмар Дженкина. В связи с этим совершенно парадоксален тот факт, что первой реакцией большинства биологов на открытие генов было мнение, что генетика опровергает теорию Дарвина, хотя при этом никто из серьезных ученых не отвергал реальность эволюции. Основной причиной кажущейся несовместимости дарвинизма и генетики было то, что основатели генетики, в частности Хуго де Фриз, наиболее плодотворный ученый из трех биологов, переоткрывших законы Менделя, рассматривали мутации генов как прерывистые, скачкообразные наследственные изменения, противоречащие постепенной эволюции в теории Дарвина. Мутации с малым фенотипическим эффектом считались неотъемлемой чертой дарвинизма, в полном соответствии с «Происхождением…». Поэтому де Фриз полагал, что его теория мутаций «антидарвинистская». Таким образом, столетний юбилей Дарвина, а также 50-летие публикации «Происхождения…» в 1909 году были далеко не триумфальными, даже на фоне резкого роста генетических исследований и введения термина «ген» Вильгельмом Йогансеном в том же году.
Еще удивительнее, что сам Мендель, очевидно хорошо знакомый с «Происхождением…»[7], не рассматривал свое открытие в контексте теории Дарвина. Ожидать установления этой жизненно важной связи пришлось не только до возрождения генетики на заре XX века, но также до появления популяционной генетики в 1920-х годах. Повторное открытие механизма наследования и рождение генетики дало мощный толчок развитию дарвинизма, так как выявление дискретных носителей наследственности устраняло кошмар Дженкина. В связи с этим совершенно парадоксален тот факт, что первой реакцией большинства биологов на открытие генов было мнение, что генетика опровергает теорию Дарвина, хотя при этом никто из серьезных ученых не отвергал реальность эволюции. Основной причиной кажущейся несовместимости дарвинизма и генетики было то, что основатели генетики, в частности Хуго де Фриз, наиболее плодотворный ученый из трех биологов, переоткрывших законы Менделя, рассматривали мутации генов как прерывистые, скачкообразные наследственные изменения, противоречащие постепенной эволюции в теории Дарвина. Мутации с малым фенотипическим эффектом считались неотъемлемой чертой дарвинизма, в полном соответствии с «Происхождением…». Поэтому де Фриз полагал, что его теория мутаций «антидарвинистская». Таким образом, столетний юбилей Дарвина, а также 50-летие публикации «Происхождения…» в 1909 году были далеко не триумфальными, даже на фоне резкого роста генетических исследований и введения термина «ген» Вильгельмом Йогансеном в том же году.
Популяционная генетика, теорема Фишера, адаптивные ландшафты, генетический дрейф и «эволюционная тяга»
Основы крайне важного синтеза дарвинизма и генетики были заложены в конце 1920-х – начале 1930-х годов тремя выдающимися генетиками-теоретиками – Рональдом Фишером, Сьюэлом Райтом и Дж. Б. С. Холдейном. Основываясь на точных математических и статистических расчетах, они создали идеализированную модель эволюции в биологической популяции. Вероятно, великий ученый-статистик Фишер первым обратил внимание, что генетика никоим образом не противоречит дарвинизму, а, напротив, предоставляет естественный и твердый фундамент для теории дарвиновской эволюции. Фишер обобщил свои выводы в исторической работе 1930 года «Генетическая теория естественного отбора» (Fisher, 1930), пожалуй, втором по значимости для эволюционной биологии труде после дарвиновского «Происхождения…»[8]. Это стало началом блистательного возрождения дарвинизма, позже получившего название современный синтез (термин, используемый преимущественно в США), или неодарвинизм (в британской и европейских традициях)[9].
Нет ни надобности, ни практической возможности излагать здесь основы популяционной генетики[10]. Можно, однако, лаконично представить некоторые обобщения, имеющие отношение к остальной части обсуждения современной эволюционной биологии. Пусть и поверхностное, но такое резюме здесь будет существенно. По сути, основатели популяционной генетики осознали простой факт, что эволюция не действует на изолированные организмы или абстрактные виды, а направлена на конкретные группы скрещивающихся особей, называемые популяциями. Размер и структура эволюционирующей популяции в большой степени определяют направление и результат эволюции. В частности, Фишер сформулировал и доказал фундаментальную теорему естественного отбора (известную как теорема Фишера), в которой утверждается, что интенсивность отбора (и, следовательно, скорость эволюции путем отбора) пропорциональна величине генетической дисперсии по приспособленности эволюционирующей популяции, которая, в свою очередь, пропорциональна эффективному размеру популяции.
В табл. 1–1 собраны основные определения и уравнения, описывающие эффекты мутаций и давления отбора на устранение или закрепление мутантных аллелей в зависимости от эффективного размера популяции. Качественная суть этих уравнений в том, что при одинаковой скорости мутаций в популяции большего эффективного размера отбор более интенсивный. В таких популяциях даже мутации с небольшим положительным коэффициентом отбора («слегка» благоприятные мутации) закрепляются быстро. С другой стороны, мутации даже с очень маленьким отрицательным коэффициентом селекции («слегка» вредные мутации) быстро устраняются. Данный эффект был строго сформулирован в теореме Фишера.
Таблица 1–1. Фундаментальное соотношение, описывающее роль отбора и генетический дрейф в эволюции популяции

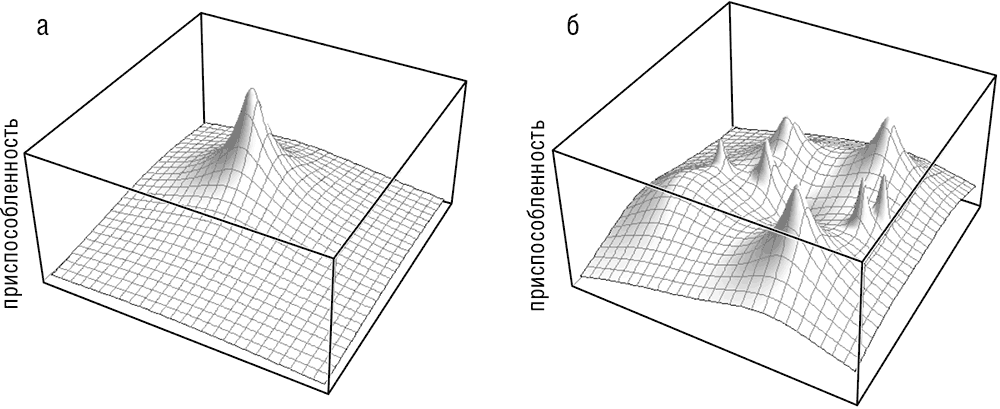
Из теоремы Фишера следует, что при эволюции, направляемой только естественным отбором, средняя приспособленность популяции не может уменьшаться (если, конечно, популяция собирается выжить). Пожалуй, наилучшим образом это можно представить с помощью образа «адаптивного ландшафта», который впервые был предложен другим отцом-основателем популяционной генетики, Сьюэлом Райтом. Райт создал этот чрезвычайно удачный образ в ответ на просьбу своего научного руководителя представить результаты математического анализа отбора в приемлемой для биологов форме. Благодаря своей простоте и изяществу это представление адаптивной эволюции сохраняет свою ценность по сей день и стимулировало многочисленные исследования, в результате которых появились более сложные и менее интуитивно понятные адаптивные ландшафты, в том числе и многомерные (Gavrilets, 2004)[11]. В соответствии с теоремой Фишера популяция, эволюция которой идет только за счет отбора (строго говоря, популяция бесконечного размера – такие популяции, естественно, не существуют, но являются удобной абстракцией, часто используемой в популяционной генетике), никогда не будет двигаться вниз по адаптивному ландшафту (см. рис. 1–1). Легко представить, что адаптивный ландшафт, как и обычный ландшафт, может иметь самую различную форму. При определенных обстоятельствах ландшафт может быть очень гладким, с единственным пиком, соответствующим глобальному адаптивному максимуму (иногда такой ландшафт образно называют «гора Фудзияма» (см. рис. 1–1а). Реальный ландшафт, однако, неровный и содержит многочисленные пики различной высоты, разделенные долинами (см. рис. 1–1б). Формально, согласно теореме Фишера (и в целом, в соответствии с теорией Дарвина), популяция, эволюционирующая с помощью отбора, может только подниматься вверх и, таким образом, достигнуть только локального пика, даже если его высота значительно меньше, чем высота глобального пика (см. рис. 1–1а). Теория Дарвина и СТЭ утверждают, что движение популяции через долины запрещено, так как неизбежно подразумевает фазу спуска. Однако развитие популяционной генетики и ее применение к эволюционным процессам изменило эту упорядоченную картину, привнеся в нее понятие «дрейфа генов», ключевую идею эволюционной биологии, которую также предложил Райт.
Рис. 1–1. Адаптивные ландшафты: а – «гора Фудзияма» с единственным (глобальным) пиком; б – «пересеченная местность» неровного адаптивного ландшафта
Как подчеркивалось ранее, Дарвин признавал важную роль случайности в эволюции, но эта роль была ограничена только одной частью эволюционного процесса: появлением изменений (в современной терминологии – мутаций). В остальном эволюция рассматривалась как строго детерминистский процесс, где отбором закрепляются выгодные мутации, а все прочие мутации устраняются без какого-либо вреда для дальнейшего существования популяции. Однако при рассмотрении популяции в динамике картина значительно меняется. Основатели количественной популяционной генетики отразили в простых формулах зависимость интенсивности отбора от размера популяции и частоты мутаций (см. табл. 1–1 и рис. 1–2). Отбор эффективен в большой популяции, и мутация, несущая незначительное преимущество, почти наверняка закрепится (в популяции бесконечного размера закрепляется мутация с бесконечно малым положительным коэффициентом отбора). Райт понял, что в малой популяции, особенно при низкой частоте мутаций, эволюционный процесс идет по-другому. В такой популяции решающую роль играет дрейф генов, с помощью которого случайным образом часто закрепляются нейтральные и даже вредные (но, конечно, не летальные) мутации. Очевидно, с помощью генетического дрейфа эволюционирующая популяция может избежать однонаправленного подъема по адаптивному ландшафту и может спускаться (см. рис. 1–2)[12]. Преимущественно это выражается в движении вниз и последующем вымирании, однако если долина, отделяющая один локальный пик от другого, возможно даже более высокого, достаточно узкая, становится возможным переход через нее и последующее восхождение на более высокую вершину (см. рис. 1–2). Введение понятия генетического дрейфа в изучение эволюции является центральным в моем рассказе. Это новый уровень проявления случая. Хотя Дарвин и его ближайшие последователи видели роль случая в появлении наследуемых изменений (мутаций), дрейф вводит случайность на следующей стадии, то есть при закреплении этих изменений, забирая у отбора часть ответственности. В этой книге я исследую, насколько значимой может быть роль дрейфа в различных ситуациях в ходе эволюции.
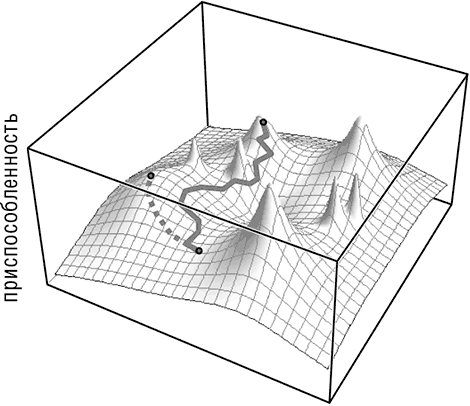 Рис. 1–2. Эволюционные траектории на неровном адаптивном ландшафте. Пунктирной линией обозначается эволюционная траектория при высоком значении эффективного размера популяции. Сплошной линией обозначается эволюционная траектория при низком значении эффективного размера популяции.
Рис. 1–2. Эволюционные траектории на неровном адаптивном ландшафте. Пунктирной линией обозначается эволюционная траектория при высоком значении эффективного размера популяции. Сплошной линией обозначается эволюционная траектория при низком значении эффективного размера популяции.
Джон Мейнард Смит и, позднее, Джон Гиллеспи разработали теорию и компьютерные модели для демонстрации существования особого режима нейтральной эволюции, который слабо зависит от эффективного размера популяции и актуален даже в популяции бесконечного размера с сильным отбором. Этот способ нейтрального закрепления мутаций стал известен как «генетическая тяга» и относится к ситуациям, в которых одна или несколько нейтральных или даже умеренно вредных мутаций распространяются в популяции и в конечном итоге закрепляются, будучи связанными с полезной мутацией. Иными словами, нейтральные или вредные аллели «двигаются в одной повозке» вместе с полезным аллелем (Barton, 2000). Похоже, что некоторые данные и модели популяционной генетики свидетельствуют, что «движение в одной повозке» даже важнее для эволюции популяции с половым размножением, чем дрейф. Очевидно, что эффект «езды в одной повозке» обусловлен совокупным воздействием естественного отбора и нейтральной изменчивостью в различных участках генома и, в отличие от дрейфа, может происходить даже в популяции бесконечно большого эффективного размера (Gillespie, 2000).
За счет эффекта «движения в одной повозке» даже в больших популяциях могут закрепляться умеренно вредные мутации, что, соответственно, дает этой популяции возможность пересекать долины адаптивного ландшафта.
Нет ни надобности, ни практической возможности излагать здесь основы популяционной генетики[10]. Можно, однако, лаконично представить некоторые обобщения, имеющие отношение к остальной части обсуждения современной эволюционной биологии. Пусть и поверхностное, но такое резюме здесь будет существенно. По сути, основатели популяционной генетики осознали простой факт, что эволюция не действует на изолированные организмы или абстрактные виды, а направлена на конкретные группы скрещивающихся особей, называемые популяциями. Размер и структура эволюционирующей популяции в большой степени определяют направление и результат эволюции. В частности, Фишер сформулировал и доказал фундаментальную теорему естественного отбора (известную как теорема Фишера), в которой утверждается, что интенсивность отбора (и, следовательно, скорость эволюции путем отбора) пропорциональна величине генетической дисперсии по приспособленности эволюционирующей популяции, которая, в свою очередь, пропорциональна эффективному размеру популяции.
В табл. 1–1 собраны основные определения и уравнения, описывающие эффекты мутаций и давления отбора на устранение или закрепление мутантных аллелей в зависимости от эффективного размера популяции. Качественная суть этих уравнений в том, что при одинаковой скорости мутаций в популяции большего эффективного размера отбор более интенсивный. В таких популяциях даже мутации с небольшим положительным коэффициентом отбора («слегка» благоприятные мутации) закрепляются быстро. С другой стороны, мутации даже с очень маленьким отрицательным коэффициентом селекции («слегка» вредные мутации) быстро устраняются. Данный эффект был строго сформулирован в теореме Фишера.
Таблица 1–1. Фундаментальное соотношение, описывающее роль отбора и генетический дрейф в эволюции популяции
Из теоремы Фишера следует, что при эволюции, направляемой только естественным отбором, средняя приспособленность популяции не может уменьшаться (если, конечно, популяция собирается выжить). Пожалуй, наилучшим образом это можно представить с помощью образа «адаптивного ландшафта», который впервые был предложен другим отцом-основателем популяционной генетики, Сьюэлом Райтом. Райт создал этот чрезвычайно удачный образ в ответ на просьбу своего научного руководителя представить результаты математического анализа отбора в приемлемой для биологов форме. Благодаря своей простоте и изяществу это представление адаптивной эволюции сохраняет свою ценность по сей день и стимулировало многочисленные исследования, в результате которых появились более сложные и менее интуитивно понятные адаптивные ландшафты, в том числе и многомерные (Gavrilets, 2004)[11]. В соответствии с теоремой Фишера популяция, эволюция которой идет только за счет отбора (строго говоря, популяция бесконечного размера – такие популяции, естественно, не существуют, но являются удобной абстракцией, часто используемой в популяционной генетике), никогда не будет двигаться вниз по адаптивному ландшафту (см. рис. 1–1). Легко представить, что адаптивный ландшафт, как и обычный ландшафт, может иметь самую различную форму. При определенных обстоятельствах ландшафт может быть очень гладким, с единственным пиком, соответствующим глобальному адаптивному максимуму (иногда такой ландшафт образно называют «гора Фудзияма» (см. рис. 1–1а). Реальный ландшафт, однако, неровный и содержит многочисленные пики различной высоты, разделенные долинами (см. рис. 1–1б). Формально, согласно теореме Фишера (и в целом, в соответствии с теорией Дарвина), популяция, эволюционирующая с помощью отбора, может только подниматься вверх и, таким образом, достигнуть только локального пика, даже если его высота значительно меньше, чем высота глобального пика (см. рис. 1–1а). Теория Дарвина и СТЭ утверждают, что движение популяции через долины запрещено, так как неизбежно подразумевает фазу спуска. Однако развитие популяционной генетики и ее применение к эволюционным процессам изменило эту упорядоченную картину, привнеся в нее понятие «дрейфа генов», ключевую идею эволюционной биологии, которую также предложил Райт.
Рис. 1–1. Адаптивные ландшафты: а – «гора Фудзияма» с единственным (глобальным) пиком; б – «пересеченная местность» неровного адаптивного ландшафта
Как подчеркивалось ранее, Дарвин признавал важную роль случайности в эволюции, но эта роль была ограничена только одной частью эволюционного процесса: появлением изменений (в современной терминологии – мутаций). В остальном эволюция рассматривалась как строго детерминистский процесс, где отбором закрепляются выгодные мутации, а все прочие мутации устраняются без какого-либо вреда для дальнейшего существования популяции. Однако при рассмотрении популяции в динамике картина значительно меняется. Основатели количественной популяционной генетики отразили в простых формулах зависимость интенсивности отбора от размера популяции и частоты мутаций (см. табл. 1–1 и рис. 1–2). Отбор эффективен в большой популяции, и мутация, несущая незначительное преимущество, почти наверняка закрепится (в популяции бесконечного размера закрепляется мутация с бесконечно малым положительным коэффициентом отбора). Райт понял, что в малой популяции, особенно при низкой частоте мутаций, эволюционный процесс идет по-другому. В такой популяции решающую роль играет дрейф генов, с помощью которого случайным образом часто закрепляются нейтральные и даже вредные (но, конечно, не летальные) мутации. Очевидно, с помощью генетического дрейфа эволюционирующая популяция может избежать однонаправленного подъема по адаптивному ландшафту и может спускаться (см. рис. 1–2)[12]. Преимущественно это выражается в движении вниз и последующем вымирании, однако если долина, отделяющая один локальный пик от другого, возможно даже более высокого, достаточно узкая, становится возможным переход через нее и последующее восхождение на более высокую вершину (см. рис. 1–2). Введение понятия генетического дрейфа в изучение эволюции является центральным в моем рассказе. Это новый уровень проявления случая. Хотя Дарвин и его ближайшие последователи видели роль случая в появлении наследуемых изменений (мутаций), дрейф вводит случайность на следующей стадии, то есть при закреплении этих изменений, забирая у отбора часть ответственности. В этой книге я исследую, насколько значимой может быть роль дрейфа в различных ситуациях в ходе эволюции.
Джон Мейнард Смит и, позднее, Джон Гиллеспи разработали теорию и компьютерные модели для демонстрации существования особого режима нейтральной эволюции, который слабо зависит от эффективного размера популяции и актуален даже в популяции бесконечного размера с сильным отбором. Этот способ нейтрального закрепления мутаций стал известен как «генетическая тяга» и относится к ситуациям, в которых одна или несколько нейтральных или даже умеренно вредных мутаций распространяются в популяции и в конечном итоге закрепляются, будучи связанными с полезной мутацией. Иными словами, нейтральные или вредные аллели «двигаются в одной повозке» вместе с полезным аллелем (Barton, 2000). Похоже, что некоторые данные и модели популяционной генетики свидетельствуют, что «движение в одной повозке» даже важнее для эволюции популяции с половым размножением, чем дрейф. Очевидно, что эффект «езды в одной повозке» обусловлен совокупным воздействием естественного отбора и нейтральной изменчивостью в различных участках генома и, в отличие от дрейфа, может происходить даже в популяции бесконечно большого эффективного размера (Gillespie, 2000).
За счет эффекта «движения в одной повозке» даже в больших популяциях могут закрепляться умеренно вредные мутации, что, соответственно, дает этой популяции возможность пересекать долины адаптивного ландшафта.
Положительный и очищающий (отрицательный) отбор: классификация форм отбора
Дарвин думал о естественном отборе в первую очередь с точки зрения закрепления благоприятных изменений. Он понимал, что эволюцией отсеиваются вредные изменения, но не интерпретировал эту ликвидацию в одной плоскости с естественным отбором. С развитием СТЭ понятие отбора было расширено за счет включения «очищающего» (отрицательного) отбора, который в некоторых фазах эволюции оказывается более распространенным (на самом деле на порядок более распространенным), чем «дарвиновский» положительный отбор. По сути, очищающий отбор – это просто элиминация неприспособленных особей. Тем не менее выделение этого процесса в особую форму отбора представляется оправданным и важным, потому что оно подчеркивает ключевую роль элиминации в формировании (сдерживании) биологического разнообразия на всех уровнях. Проще говоря, изменение допускается, только если оно не наносит существенного вреда никаким из выживающих особей. Интересен и открыт вопрос, до какой степени эти ограничения фактически сужают пространство, доступное для эволюции, и я коснусь этого вопроса позднее (см., в частности, гл. 3, 8 и 9).