Тонкая, но важная разница существует между очищающим отбором и стабилизирующим отбором, который является еще одной из форм отбора, которая действует на распределение частот отличительных признаков. Таким образом, выделяются следующие формы отбора: стабилизирующий отбор, основанный в первую очередь на очищающем отборе, движущий отбор, обусловленный положительным (дарвиновским) отбором, и более экзотические режимы дизруптивного и балансирующего отбора, которые являются результатом сочетания многочисленных ограничений (см. рис. 1–3).
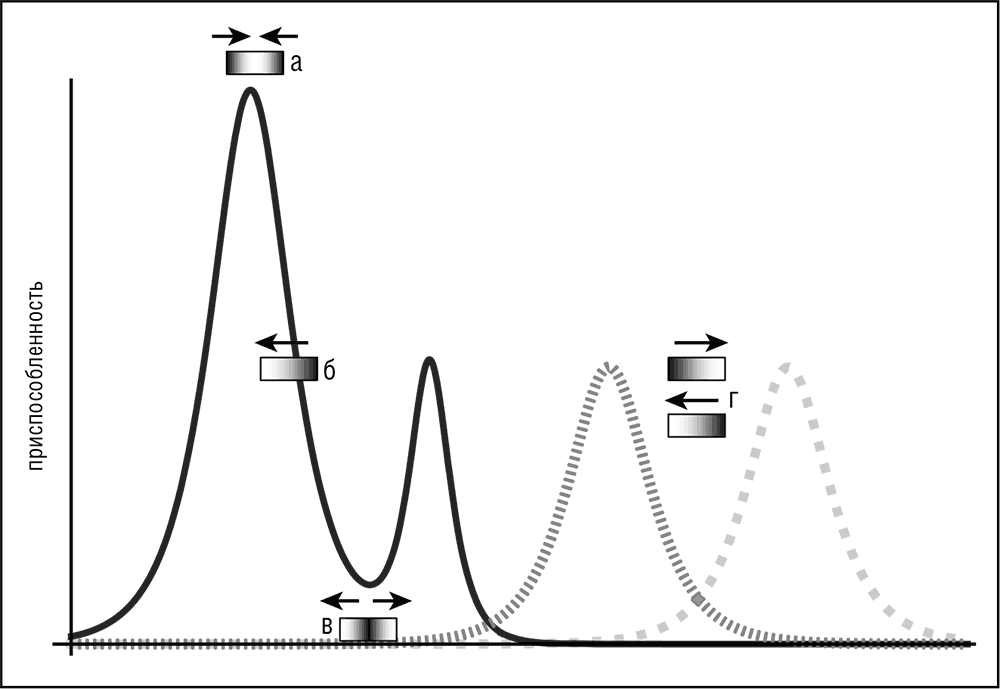 Рис. 1–3. Четыре различные формы отбора в эволюционирующей популяции: а — стабилизирующий отбор (адаптивный ландшафт представлен сплошной линией); б — движущий отбор (адаптивный ландшафт представлен сплошной линией); в — дизруптивный отбор (адаптивный ландшафт представлен сплошной линией); г — балансирующий отбор (адаптивный ландшафт периодически меняется, переключаясь между двумя пунктирными линиями)
Рис. 1–3. Четыре различные формы отбора в эволюционирующей популяции: а — стабилизирующий отбор (адаптивный ландшафт представлен сплошной линией); б — движущий отбор (адаптивный ландшафт представлен сплошной линией); в — дизруптивный отбор (адаптивный ландшафт представлен сплошной линией); г — балансирующий отбор (адаптивный ландшафт периодически меняется, переключаясь между двумя пунктирными линиями)
Синтетическая теория эволюции
Краткий обзор главы
Рекомендуемая дополнительная литература
Глава 2. От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Репликация цифровых носителей информации: центральный принцип биологии и необходимое и достаточное условие эволюции
Синтетическая теория эволюции
Объединение дарвиновской теории эволюции и генетики, состоявшееся в основополагающих исследованиях Фишера, Райта и Холдейна, подготовило почву для рождения синтетической теории эволюционной биологии. Само название идет от одноименной книги, опубликованной Джулианом Хаксли в 1942 году (Huxley, 2010), однако концептуальная структура СТЭ полностью сформировалась только в 1959 году в ходе мероприятий, посвященных 100-летнему юбилею «Происхождения…». Новая синтетическая теория стала результатом работы многих выдающихся ученых. Можно утверждать, что главными архитекторами СТЭ были экспериментальный генетик Феодосий Добржанский, зоолог Эрнст Майр и палеонтолог Джордж Гейлорд Симпсон. Экспериментальные и полевые работы Добржанского с плодовой мушкой Drosophila melanogaster принесли насущные фактические свидетельства в поддержку теории популяционной генетики и стали первой крупномасштабной экспериментальной проверкой идеи естественного отбора. Книга Добржанского «Генетика и происхождение видов» (Dobzhansky, 1951) явилась основным программным документом СТЭ, в котором он сузил понятие эволюции до «изменения частоты аллеля в генетическом пуле». Знаменита также крылатая фраза Добржанского о том, что «ничто в биологии не имеет смысла, кроме как в свете эволюции»[13] (см. больше о «смысле» в прил. I). Заслугой Эрнста Майра, как никакого другого ученого, является серьезная, крайне влиятельная попытка теоретического решения принципиальной проблемы, поставленной Дарвином, – происхождения видов. Майр сформулировал так называемую биологическую концепцию вида, согласно которой видообразование происходит, когда две популяции (размножающиеся половым путем) изолированы друг от друга достаточно долго, чтобы обеспечить необратимую генетическую несовместимость (Mayr, 1963).
Симпсон реконструировал наиболее полную (на тот момент) картину эволюции жизни на основании палеонтологической летописи (Simpson, 1983). Замечательно, что Симпсон осознал стазис (отсутствие существенных изменений) в эволюции большинства видов и резкую смену доминантных видов. Он ввел понятие квантовой эволюции, которое предвосхитило теорию прерывистого равновесия, предложенную Стивеном Джеем Гулдом и Нильсом Элдриджем (см. гл. 2).
Консолидация СТЭ в 1950-х годах была довольно странным процессом, сопровождавшимся странным «затвердеванием» (выражение Гулда) основных идей Дарвина (Gould, 2002). Так, доктрина СТЭ фактически отбросила идею Райта о случайном дрейфе генов и его эволюционной важности и стала бескомпромиссно панадаптационистской. Более того, сам Симпсон отказался от идеи квантовой эволюции, так что градуализм продолжал оставаться одним из неоспоримых столпов СТЭ. Такое «затвердевание» сделало СТЭ относительно узкой, в некотором смысле даже догматичной, системой.
Чтобы продолжить обсуждение эволюции эволюционной биологии и ее преображение в век геномики, представляется необходимым кратко резюмировать основные принципы эволюции, впервые сформулированные Дарвином, затем усовершенствованные первым поколением биологов-эволюционистов и, наконец, кодифицированные в СТЭ. Мы будем возвращаться к каждому из этих ключевых моментов на протяжении всей книги.
1. Ненаправленное случайное изменение – это главный процесс, обеспечивающий материал для эволюции. Дарвин впервые показал, что случайность является основным фактором в истории жизни, и это, несомненно, было одной из его наиболее важных идей. Дарвин также признавал роль направленной, ламарковской изменчивости и в последующих изданиях «Происхождения…» склонялся даже к более весомой роли этого механизма эволюции. Однако СТЭ твердо настаивает на том, что случайные мутации являются единственным источником эволюционно значимой изменчивости.
2. Действие эволюции заключается в фиксации редких выгодных изменений и элиминации вредных изменений. Согласно Дарвину и СТЭ, в этом состоит процесс естественного отбора, который, наряду со случайной изменчивостью, является основной движущей силой эволюции. Естественный отбор, очевидно сходный и навеянный «невидимой рукой» рынка, которая, по теории Адама Смита, управляет экономикой, был первым из когда-либо предложенных механизмов эволюции, который был прост и правдоподобен и не требовал изначально мистического подхода. Таким образом, это вторая ключевая идея Дарвина. Сьюэл Райт подчеркивал, что случайность может играть вспомогательную роль не только в возникновении, но также и в закреплении изменений в ходе эволюции с помощью дрейфа генов, в результате которого случайно сохраняются нейтральные или умеренно вредные изменения. Согласно теории популяционной генетики, дрейф генов особенно значим в небольших популяциях, проходящих через «бутылочное горлышко». «Генетическая тяга», или «езда в одной повозке», – это другая форма случайного закрепления невыгодных мутаций. Однако СТЭ в ее догматизированной форме фактически отрицает стохастические процессы в эволюции, кроме возникновения изменений, и придерживается полностью адаптационистского (панадаптационистского) взгляда на эволюцию. Такая модель неизбежно приводит к концепции «прогресса», постепенного улучшения «органов» в ходе эволюции. Дарвин поддерживал эту идею как основное направление развития, несмотря на четкое понимание, что организмы все еще далеки от совершенства в плане адаптивности, как можно прекрасно увидеть на примере рудиментарных органов, и несмотря на свое резко отрицательное отношение к любым формам ламарковского внутреннего стремления к совершенству. СТЭ уходит от прогресса как антропоморфной идеи, но тем не менее поддерживает общую концепцию эволюции от простых форм к сложным.
3. Полезные изменения, закрепляемые естественным отбором, бесконечно малы (в современной терминологии, эволюционно значимые мутации обладают бесконечно малым влиянием на приспособленность), поэтому эволюция происходит путем постепенного накопления этих слабых изменений. Дарвин был убежден, что в основе его теории лежит строгий градуализм: «Естественный отбор действует только путем сохранения и кумулирования малых наследственных модификаций, каждая из которых выгодна для сохраняемого существа… Если бы возможно было показать, что существует сложный орган, который не мог образоваться путем многочисленных последовательных слабых модификаций, моя теория потерпела бы полное крушение» («Происхождение видов…», гл. 6 [цит. по: Дарвин Ч. Сочинения. Т. 3 / Пер. с англ. К. А. Тимирязева, С. Л. Соболя. M.: Изд-во АН СССР, 1939]). Даже некоторые современники Дарвина полагали, что это излишняя, искусственная строгость теории. В частности, хорошо известны заблаговременные возражения со стороны Томаса Гексли. Еще до публикации «Происхождения…» Гексли написал Дарвину: «Вы взяли на себя ненужный груз безоговорочного следования принципу Natura non facit saltum» (http://aleph0.clarku.edu/huxley/). Несмотря на эти своевременные предостережения и даже на идею Симпсона о квантовом характере эволюции, СТЭ бескомпромиссно настаивает на градуализме.
4. Униформизм (термин был заимствован Дарвином из геологии Лайеля) – это один из аспектов классической эволюционной биологии, который связан, но в то же время отличается от принципиального градуализма. Это убеждение, что эволюционные процессы не изменялись по существу на всем протяжении истории жизни.
5. Следующий ключевой принцип логически связан с градуализмом и униформизмом: макроэволюция (происхождение видов и высших таксонов) управляется теми же механизмами, что и микроэволюция (эволюция внутри вида). Главным апологетом данного принципа был Добржанский, определивший эволюцию как изменение частоты аллелей в популяциях. Дарвин не пользовался терминами микроэволюция и макроэволюция; тем не менее достаточность внутривидовых процессов для объяснения происхождения видов и, в более широком плане, всей эволюции жизни можно считать центральной аксиомой Дарвина (или, возможно, фундаментальной теоремой, такой, однако, для которой у Дарвина не было даже и намека на доказательство). Представляется разумным говорить о данном принципе как об «универсальном униформизме»: эволюционные процессы одинаковы не только на протяжении всей истории жизни, но и на разных уровнях эволюционных изменений, включая крупные преобразования. Загадка взаимосвязи между микроэволюцией и макроэволюцией является в некотором смысле осью эволюционной биологии, поэтому мы будем постоянно возвращаться к ней в этой книге.
6. Эволюцию жизни можно адекватно представить в виде «огромного дерева», что и подчеркнуто единственной иллюстрацией в «Происхождении…» (в гл. 4). Дарвин представил древо жизни только как общую идею и не пытался исследовать фактический порядок ветвления. Древо было заселено реальными формами жизни, насколько они были известны в то время, одним из главных последователей Дарвина, знаменитым немецким биологом Эрнстом Геккелем. Основатели СТЭ не проявляли большого интереса к древу жизни, но они, несомненно, включали его в теорию как описание эволюции животных и растений, убедительно поддержанное палеонтологической летописью в XX веке. Однако микробы, определяющее значение которых в глобальной экологии становилось все более очевидным, фактически остались за пределами эволюционной биологии.
7. Концепция единого древа жизни имеет следствие, которое заслуживает статуса отдельного принципа: существующее в настоящее время разнообразие форм жизни произошло от общего предка (или нескольких форм-предшественников, в соответствии с осторожной формулировкой Дарвина в главе 14 «Происхождения…», см. Darwin, 1859). Спустя много лет он был назван «последним универсальным клеточным предком» (Last Universal Cellular Ancestor, LUCA[14]). Для создателей СТЭ существование LUCA не вызывало сомнений, но они, по-видимому, не считали реалистичной или научно важной целью прояснение его природы.
Симпсон реконструировал наиболее полную (на тот момент) картину эволюции жизни на основании палеонтологической летописи (Simpson, 1983). Замечательно, что Симпсон осознал стазис (отсутствие существенных изменений) в эволюции большинства видов и резкую смену доминантных видов. Он ввел понятие квантовой эволюции, которое предвосхитило теорию прерывистого равновесия, предложенную Стивеном Джеем Гулдом и Нильсом Элдриджем (см. гл. 2).
Консолидация СТЭ в 1950-х годах была довольно странным процессом, сопровождавшимся странным «затвердеванием» (выражение Гулда) основных идей Дарвина (Gould, 2002). Так, доктрина СТЭ фактически отбросила идею Райта о случайном дрейфе генов и его эволюционной важности и стала бескомпромиссно панадаптационистской. Более того, сам Симпсон отказался от идеи квантовой эволюции, так что градуализм продолжал оставаться одним из неоспоримых столпов СТЭ. Такое «затвердевание» сделало СТЭ относительно узкой, в некотором смысле даже догматичной, системой.
Чтобы продолжить обсуждение эволюции эволюционной биологии и ее преображение в век геномики, представляется необходимым кратко резюмировать основные принципы эволюции, впервые сформулированные Дарвином, затем усовершенствованные первым поколением биологов-эволюционистов и, наконец, кодифицированные в СТЭ. Мы будем возвращаться к каждому из этих ключевых моментов на протяжении всей книги.
1. Ненаправленное случайное изменение – это главный процесс, обеспечивающий материал для эволюции. Дарвин впервые показал, что случайность является основным фактором в истории жизни, и это, несомненно, было одной из его наиболее важных идей. Дарвин также признавал роль направленной, ламарковской изменчивости и в последующих изданиях «Происхождения…» склонялся даже к более весомой роли этого механизма эволюции. Однако СТЭ твердо настаивает на том, что случайные мутации являются единственным источником эволюционно значимой изменчивости.
2. Действие эволюции заключается в фиксации редких выгодных изменений и элиминации вредных изменений. Согласно Дарвину и СТЭ, в этом состоит процесс естественного отбора, который, наряду со случайной изменчивостью, является основной движущей силой эволюции. Естественный отбор, очевидно сходный и навеянный «невидимой рукой» рынка, которая, по теории Адама Смита, управляет экономикой, был первым из когда-либо предложенных механизмов эволюции, который был прост и правдоподобен и не требовал изначально мистического подхода. Таким образом, это вторая ключевая идея Дарвина. Сьюэл Райт подчеркивал, что случайность может играть вспомогательную роль не только в возникновении, но также и в закреплении изменений в ходе эволюции с помощью дрейфа генов, в результате которого случайно сохраняются нейтральные или умеренно вредные изменения. Согласно теории популяционной генетики, дрейф генов особенно значим в небольших популяциях, проходящих через «бутылочное горлышко». «Генетическая тяга», или «езда в одной повозке», – это другая форма случайного закрепления невыгодных мутаций. Однако СТЭ в ее догматизированной форме фактически отрицает стохастические процессы в эволюции, кроме возникновения изменений, и придерживается полностью адаптационистского (панадаптационистского) взгляда на эволюцию. Такая модель неизбежно приводит к концепции «прогресса», постепенного улучшения «органов» в ходе эволюции. Дарвин поддерживал эту идею как основное направление развития, несмотря на четкое понимание, что организмы все еще далеки от совершенства в плане адаптивности, как можно прекрасно увидеть на примере рудиментарных органов, и несмотря на свое резко отрицательное отношение к любым формам ламарковского внутреннего стремления к совершенству. СТЭ уходит от прогресса как антропоморфной идеи, но тем не менее поддерживает общую концепцию эволюции от простых форм к сложным.
3. Полезные изменения, закрепляемые естественным отбором, бесконечно малы (в современной терминологии, эволюционно значимые мутации обладают бесконечно малым влиянием на приспособленность), поэтому эволюция происходит путем постепенного накопления этих слабых изменений. Дарвин был убежден, что в основе его теории лежит строгий градуализм: «Естественный отбор действует только путем сохранения и кумулирования малых наследственных модификаций, каждая из которых выгодна для сохраняемого существа… Если бы возможно было показать, что существует сложный орган, который не мог образоваться путем многочисленных последовательных слабых модификаций, моя теория потерпела бы полное крушение» («Происхождение видов…», гл. 6 [цит. по: Дарвин Ч. Сочинения. Т. 3 / Пер. с англ. К. А. Тимирязева, С. Л. Соболя. M.: Изд-во АН СССР, 1939]). Даже некоторые современники Дарвина полагали, что это излишняя, искусственная строгость теории. В частности, хорошо известны заблаговременные возражения со стороны Томаса Гексли. Еще до публикации «Происхождения…» Гексли написал Дарвину: «Вы взяли на себя ненужный груз безоговорочного следования принципу Natura non facit saltum» (http://aleph0.clarku.edu/huxley/). Несмотря на эти своевременные предостережения и даже на идею Симпсона о квантовом характере эволюции, СТЭ бескомпромиссно настаивает на градуализме.
4. Униформизм (термин был заимствован Дарвином из геологии Лайеля) – это один из аспектов классической эволюционной биологии, который связан, но в то же время отличается от принципиального градуализма. Это убеждение, что эволюционные процессы не изменялись по существу на всем протяжении истории жизни.
5. Следующий ключевой принцип логически связан с градуализмом и униформизмом: макроэволюция (происхождение видов и высших таксонов) управляется теми же механизмами, что и микроэволюция (эволюция внутри вида). Главным апологетом данного принципа был Добржанский, определивший эволюцию как изменение частоты аллелей в популяциях. Дарвин не пользовался терминами микроэволюция и макроэволюция; тем не менее достаточность внутривидовых процессов для объяснения происхождения видов и, в более широком плане, всей эволюции жизни можно считать центральной аксиомой Дарвина (или, возможно, фундаментальной теоремой, такой, однако, для которой у Дарвина не было даже и намека на доказательство). Представляется разумным говорить о данном принципе как об «универсальном униформизме»: эволюционные процессы одинаковы не только на протяжении всей истории жизни, но и на разных уровнях эволюционных изменений, включая крупные преобразования. Загадка взаимосвязи между микроэволюцией и макроэволюцией является в некотором смысле осью эволюционной биологии, поэтому мы будем постоянно возвращаться к ней в этой книге.
6. Эволюцию жизни можно адекватно представить в виде «огромного дерева», что и подчеркнуто единственной иллюстрацией в «Происхождении…» (в гл. 4). Дарвин представил древо жизни только как общую идею и не пытался исследовать фактический порядок ветвления. Древо было заселено реальными формами жизни, насколько они были известны в то время, одним из главных последователей Дарвина, знаменитым немецким биологом Эрнстом Геккелем. Основатели СТЭ не проявляли большого интереса к древу жизни, но они, несомненно, включали его в теорию как описание эволюции животных и растений, убедительно поддержанное палеонтологической летописью в XX веке. Однако микробы, определяющее значение которых в глобальной экологии становилось все более очевидным, фактически остались за пределами эволюционной биологии.
7. Концепция единого древа жизни имеет следствие, которое заслуживает статуса отдельного принципа: существующее в настоящее время разнообразие форм жизни произошло от общего предка (или нескольких форм-предшественников, в соответствии с осторожной формулировкой Дарвина в главе 14 «Происхождения…», см. Darwin, 1859). Спустя много лет он был назван «последним универсальным клеточным предком» (Last Universal Cellular Ancestor, LUCA[14]). Для создателей СТЭ существование LUCA не вызывало сомнений, но они, по-видимому, не считали реалистичной или научно важной целью прояснение его природы.
Краткий обзор главы
В своей книге «Происхождение видов…» Чарльз Дарвин тщательно собрал доказательства изменений во времени, которые охватывают мир живых существ, и впервые предложил убедительный механизм эволюции: естественный отбор. Эволюция путем естественного отбора, безусловно, является одной из самых существенных концепций, когда-либо разработанных учеными, и даже была объявлена самой важной идеей в истории человечества (Dennett, 1996). В свете этого может показаться парадоксальным, что понятие естественного отбора нередко считают простой тавтологией. Если рассуждать с точки зрения выживания наиболее приспособленных, видно, что для этого взгляда есть основания. Однако, если рассматривать весь дарвиновский сценарий эволюции в целом, его решительно не тавтологические и не тривиальные аспекты становятся очевидными. В действительности Дарвин предложил механизм преобразования случайных изменений в отнюдь не случайные адаптации, вплоть до сложнейших приспособлений, исполняющих узкоспециализированные функции и тем самым повышающих приспособленность их носителей. Если рассматривать этот процесс в терминах физики и несколько вольно следовать идеям знаменитой книги Эрвина Шредингера, дарвиновская эволюция является машиной для создания отрицательной энтропии, другими словами, порядка из беспорядка. На мой взгляд, самым главным прозрением Дарвина было осознание того, что простой механизм, лишенный какого-либо телеологического содержания, вероятно, мог только благодаря случайным изменениям привести к появлению удивительного разнообразия форм жизни, каждая из которых в совершенстве приспособлена к условиям среды своего обитания. С этой точки зрения, «невидимая рука» естественного отбора кажется почти чудесно всесильной, и нельзя не задаваться вопросом, действительно ли этого достаточно, чтобы объяснить историю жизни. Этот вопрос неоднократно использовался в качестве риторического приема креационистами всех мастей, но он также всерьез поднимался биологами-эволюционистами. В остальной части этой книги мы увидим, что разные ученые дают на него разные ответы, которые зависят от того, о каких именно ситуациях и этапах эволюции жизни идет речь.
Конечно, дарвинизм в его изначальном виде столкнулся с более значительными и непосредственными проблемами, чем вопрос о достаточности естественного отбора: Дарвин и его ранние последователи не имели представления о механизмах наследования и о том, будут ли когда-либо открыты механизмы, согласующиеся со сценарием Дарвина. В этом смысле здание теории Дарвина висело в воздухе. Повторное открытие законов генетики в начале XX века и последующее развитие теоретической и экспериментальной популяционной генетики обеспечило твердое основание для дарвиновской теории эволюции. Было показано, что, без сомнения, популяции эволюционируют посредством процесса, в котором дарвиновский естественный отбор играет важнейшую роль. СТЭ в эволюционной биологии явилась завершением работы Дарвина, последовательно объединив дарвинизм и генетику. По мере развития СТЭ заметно «окостенела», настаивая на градуализме, униформизме и, что наиболее важно, монополии естественного отбора как единственном пути эволюции. В соответствии с СТЭ все изменения, закрепляемые в ходе эволюции, являются адаптивными, по крайней мере изначально. При всех своих выдающихся достоинствах СТЭ представляет собой довольно догматичную и удручающе незаконченную теорию. Назовем три наиболее бросающиеся в глаза проблемы: СТЭ совершенно бездоказательно распространяет механизмы и закономерности, принятые в микроэволюции, на макроэволюционные процессы; она ничего не говорит об эволюции микробов, являющихся наиболее широко распространенными и многообразными формами жизни на Земле; и она даже не пытается обратиться к вопросу о происхождении жизни.
Конечно, дарвинизм в его изначальном виде столкнулся с более значительными и непосредственными проблемами, чем вопрос о достаточности естественного отбора: Дарвин и его ранние последователи не имели представления о механизмах наследования и о том, будут ли когда-либо открыты механизмы, согласующиеся со сценарием Дарвина. В этом смысле здание теории Дарвина висело в воздухе. Повторное открытие законов генетики в начале XX века и последующее развитие теоретической и экспериментальной популяционной генетики обеспечило твердое основание для дарвиновской теории эволюции. Было показано, что, без сомнения, популяции эволюционируют посредством процесса, в котором дарвиновский естественный отбор играет важнейшую роль. СТЭ в эволюционной биологии явилась завершением работы Дарвина, последовательно объединив дарвинизм и генетику. По мере развития СТЭ заметно «окостенела», настаивая на градуализме, униформизме и, что наиболее важно, монополии естественного отбора как единственном пути эволюции. В соответствии с СТЭ все изменения, закрепляемые в ходе эволюции, являются адаптивными, по крайней мере изначально. При всех своих выдающихся достоинствах СТЭ представляет собой довольно догматичную и удручающе незаконченную теорию. Назовем три наиболее бросающиеся в глаза проблемы: СТЭ совершенно бездоказательно распространяет механизмы и закономерности, принятые в микроэволюции, на макроэволюционные процессы; она ничего не говорит об эволюции микробов, являющихся наиболее широко распространенными и многообразными формами жизни на Земле; и она даже не пытается обратиться к вопросу о происхождении жизни.
Рекомендуемая дополнительная литература
Futuyma, Douglas. (2009) Evolution, 2d edition. Sunderland, MA: Sinauer Associates.
Возможно, лучший из учебников по эволюционной биологии для студентов.
Gould, Stephen Jay. (2002) The Structure of Evolutionary Theory. Cambridge, MA: Harvard University Press.
Почти 1500-страничный том, очевидно, не для слабых духом, и не многие прочитают его полностью. Тем не менее по крайней мере первая часть ценна своим четким и точным описанием истории эволюционной биологии и острой критикой СТЭ.
Hartl, Daniel L., and Andrew G. Clark. (2006) Principles of Population Genetics, 4th edition. Sunderland, MA: Sinauer Associates.
Превосходный, достаточно серьезный, но доступный для понимания учебник по популяционной генетике.
Mayr, Ernst. (2002) What Evolution Is. New York: Basic Books.
Упрощенное, но ясное и полезное представление классической эволюционной биологии одним из основателей СТЭ.
Schroedinger, Erwin. (1992) What Is Life?: With «Mind and Matter» and «Autobiographical Sketches». Cambridge, MA: Cambridge University Press. (Перевод: Шредингер Э. Что такое жизнь? Физический аспект живой клетки / Пер. с англ. 3-е изд. Ижевск: РХД, 2002.)
Первое издание этой блестящей книги вышло в 1944 году на основе лекций Шредингера (одного из основателей квантовой механики), прочитанных им в Эдинбурге, где он жил во время Второй мировой войны. Эта очевидно устаревшая, однако удивительно доходчивая, пророческая книга все еще важна для обсуждения роли энтропии и информации в биологии.
Возможно, лучший из учебников по эволюционной биологии для студентов.
Gould, Stephen Jay. (2002) The Structure of Evolutionary Theory. Cambridge, MA: Harvard University Press.
Почти 1500-страничный том, очевидно, не для слабых духом, и не многие прочитают его полностью. Тем не менее по крайней мере первая часть ценна своим четким и точным описанием истории эволюционной биологии и острой критикой СТЭ.
Hartl, Daniel L., and Andrew G. Clark. (2006) Principles of Population Genetics, 4th edition. Sunderland, MA: Sinauer Associates.
Превосходный, достаточно серьезный, но доступный для понимания учебник по популяционной генетике.
Mayr, Ernst. (2002) What Evolution Is. New York: Basic Books.
Упрощенное, но ясное и полезное представление классической эволюционной биологии одним из основателей СТЭ.
Schroedinger, Erwin. (1992) What Is Life?: With «Mind and Matter» and «Autobiographical Sketches». Cambridge, MA: Cambridge University Press. (Перевод: Шредингер Э. Что такое жизнь? Физический аспект живой клетки / Пер. с англ. 3-е изд. Ижевск: РХД, 2002.)
Первое издание этой блестящей книги вышло в 1944 году на основе лекций Шредингера (одного из основателей квантовой механики), прочитанных им в Эдинбурге, где он жил во время Второй мировой войны. Эта очевидно устаревшая, однако удивительно доходчивая, пророческая книга все еще важна для обсуждения роли энтропии и информации в биологии.
Глава 2. От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
В этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись предшественниками синтетической теории эволюции (СТЭ). На самом деле они возникали параллельно с развитием СТЭ, но были отвергнуты «каноном» вследствие «ужесточения» СТЭ. Достижения, которые обсуждаются в этой главе, относятся к интервалу между 1930 (публикация книги Рональда Фишера, которая ознаменовала вторую, зрелую стадию развития эволюционной биологии) и 1995 годами (первые сравнения полных геномов клеточных форм жизни). Моя цель здесь – вкратце обрисовать сложную сеть эволюционных идей, теорий и наблюдений, которые дополнили достаточно жесткую структуру СТЭ и стали пусковой площадкой для нового, «геномного» подхода к изучению эволюции.
Репликация цифровых носителей информации: центральный принцип биологии и необходимое и достаточное условие эволюции
Модель структуры ДНК, представленная Джеймсом Уотсоном и Фрэнсисом Криком (очевидно, основанная на рентгеновских структурах, полученных Розалинд Франклин и другими), несомненно, является одним из главных открытий не только биологии XX века, но и всей истории биологии (Watson and Crick, 1953b). Однако этот прорыв не всегда упоминается в связи с принципами биологической эволюции.
С моей точки зрения, структура ДНК и модель ее репликации, которую Уотсон и Крик описали в своей второй классической статье как непосредственное следствие структуры (Watson and Crick, 1953a), являются важнейшим фундаментальным открытием в изучении эволюции со времени публикации «Происхождения видов…». По сути, Уотсон и Крик вывели из структуры ДНК биологическое воплощение общего принципа цифрового хранения, кодирования и передачи информации. Система биологической передачи информации, которую выявили их исследования, может рассматриваться как расширение принципа машины Тьюринга, сначала через правила комплементарности нуклеотидных оснований (в процессах репликации и транскрипции), a затем в процессе трансляции, через генетический код (см. рис. 2–1). По сути, пусть и не в историческом смысле, эти открытия вытеснили концепцию Дарвина, в том смысле, что вся дарвиновская схема эволюции является прямым следствием механизма репликации ДНК. Для всех известных форм жизни биологическая передача цифровой информации влечет за собой исполнение следующих простых фундаментальных принципов[15].
Генетический материал любого организма состоит из линейной последовательности символов, четырех оснований нуклеиновых кислот, которая, прямо или косвенно, кодирует всю информацию, необходимую для построения организма[16].
Репликация генетического материала, являющегося механической основой наследственности, осуществляется на основе принципа однозначного комплементарного соответствия между A и T(U), и G и С. (Так называемые правила Чаргафа, по имени их первооткрывателя, австрийского, a затем американского химика Эрвина Чаргаффа[17].)
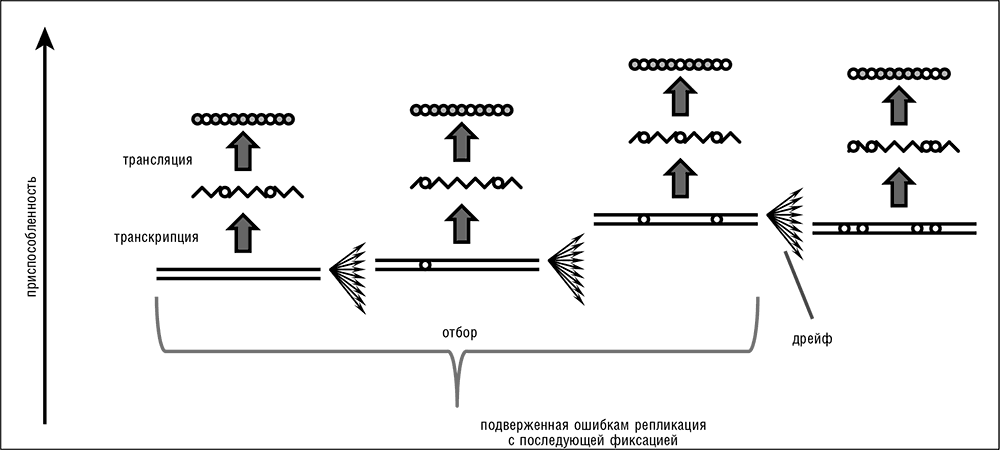 Рис. 2–1. Передача информации в биологических системах и превращение естественного отбора и генетического дрейфа в эпифеномены репликации. Белые круги на схеме обозначают изменения относительно оригинальной последовательности.
Рис. 2–1. Передача информации в биологических системах и превращение естественного отбора и генетического дрейфа в эпифеномены репликации. Белые круги на схеме обозначают изменения относительно оригинальной последовательности.
Уотсон и Крик описали эти ключевые принципы генетических систем в двух своих статьях, вышедших в 1953 году. Дальнейшие исследования добавили два очень важных аспекта:
1. Принцип комплементарности используется не только во время репликации, но и во время транскрипции ДНК во все виды РНК и во время трансляции мРНК в белок с помощью адапторных молекул тРНК.
2. Те же самые принципы цифровой репликации и декодирования применимы и для генетических систем, в которых генетический материал отличается от двойной спирали ДНК, изначально описанной Уотсоном и Криком, и состоит из РНК или односпиральной ДНК (например, у многих вирусов).
Теория информации твердо стоит на том, что передача информации абсолютно без ошибок невозможна в принципе. В реальности вероятность ошибки в любом конечном сообщении может быть сведена к минимуму, но любое снижение уровня ошибок при передаче информации возможно только за счет затраты энергии. Эта связь непосредственно следует из законов термодинамики. Центральный принцип эволюции может быть сформулирован следующим образом:
Репликация цифровых носителей информации неизбежно подвержена ошибкам, что влечет за собой эволюцию этих носителей путем естественного отбора и случайного дрейфа генов при условии, что уровень ошибок репликации ниже катастрофического порога, имеющего порядок величины от одной до десяти ошибок на геном за один цикл репликации[18].
Назовем это обобщение принципом подверженной ошибкам репликации (ПОР)[19]. Этот принцип становится самоочевидным, как только мы осознаем существование и основной механизм репликации. Он был впервые описан математически в теории Манфреда Эйгена (Eigen, 1971), который также ввел понятие концепции порога ошибки (Biebricher and Eigen, 2005) – эта теория и ее применение будут рассматриваться далее в главе 12. ПОР основывается на следующих двух предположениях, которые могут показаться очевидными, но заслуживают тем не менее особого упоминания:
С моей точки зрения, структура ДНК и модель ее репликации, которую Уотсон и Крик описали в своей второй классической статье как непосредственное следствие структуры (Watson and Crick, 1953a), являются важнейшим фундаментальным открытием в изучении эволюции со времени публикации «Происхождения видов…». По сути, Уотсон и Крик вывели из структуры ДНК биологическое воплощение общего принципа цифрового хранения, кодирования и передачи информации. Система биологической передачи информации, которую выявили их исследования, может рассматриваться как расширение принципа машины Тьюринга, сначала через правила комплементарности нуклеотидных оснований (в процессах репликации и транскрипции), a затем в процессе трансляции, через генетический код (см. рис. 2–1). По сути, пусть и не в историческом смысле, эти открытия вытеснили концепцию Дарвина, в том смысле, что вся дарвиновская схема эволюции является прямым следствием механизма репликации ДНК. Для всех известных форм жизни биологическая передача цифровой информации влечет за собой исполнение следующих простых фундаментальных принципов[15].
Генетический материал любого организма состоит из линейной последовательности символов, четырех оснований нуклеиновых кислот, которая, прямо или косвенно, кодирует всю информацию, необходимую для построения организма[16].
Репликация генетического материала, являющегося механической основой наследственности, осуществляется на основе принципа однозначного комплементарного соответствия между A и T(U), и G и С. (Так называемые правила Чаргафа, по имени их первооткрывателя, австрийского, a затем американского химика Эрвина Чаргаффа[17].)
Уотсон и Крик описали эти ключевые принципы генетических систем в двух своих статьях, вышедших в 1953 году. Дальнейшие исследования добавили два очень важных аспекта:
1. Принцип комплементарности используется не только во время репликации, но и во время транскрипции ДНК во все виды РНК и во время трансляции мРНК в белок с помощью адапторных молекул тРНК.
2. Те же самые принципы цифровой репликации и декодирования применимы и для генетических систем, в которых генетический материал отличается от двойной спирали ДНК, изначально описанной Уотсоном и Криком, и состоит из РНК или односпиральной ДНК (например, у многих вирусов).
Теория информации твердо стоит на том, что передача информации абсолютно без ошибок невозможна в принципе. В реальности вероятность ошибки в любом конечном сообщении может быть сведена к минимуму, но любое снижение уровня ошибок при передаче информации возможно только за счет затраты энергии. Эта связь непосредственно следует из законов термодинамики. Центральный принцип эволюции может быть сформулирован следующим образом:
Репликация цифровых носителей информации неизбежно подвержена ошибкам, что влечет за собой эволюцию этих носителей путем естественного отбора и случайного дрейфа генов при условии, что уровень ошибок репликации ниже катастрофического порога, имеющего порядок величины от одной до десяти ошибок на геном за один цикл репликации[18].
Назовем это обобщение принципом подверженной ошибкам репликации (ПОР)[19]. Этот принцип становится самоочевидным, как только мы осознаем существование и основной механизм репликации. Он был впервые описан математически в теории Манфреда Эйгена (Eigen, 1971), который также ввел понятие концепции порога ошибки (Biebricher and Eigen, 2005) – эта теория и ее применение будут рассматриваться далее в главе 12. ПОР основывается на следующих двух предположениях, которые могут показаться очевидными, но заслуживают тем не менее особого упоминания: