• Однако Ka и Ks положительно коррелируют между собой – таким образом, отбор затрагивает и синонимичные сайты.
• Некодирующие сайты, такие как интронные последовательности, могут использоваться как фон нейтральной эволюции при измерении отбора на синонимичных сайтах (Ks/Ki, где Ki – частота замен для интронных сайтов).
• Критерий Макдональда – Крейтмана (Aquadro, 1997; McDonald and Kreitman, 1991) широко используется для измерения отбора. Он сравнивает внутривидовые вариации (частота полиморфизма, P) с межвидовыми вариациями (дивергенция, D).
• Dn/Ds = Pn/Ps – нейтральная эволюция белковой последовательности.
• Dn/Ds < Pn/Ps – отсекающий отбор.
• Dn/Ds > Pn/Ps – положительный отбор.
Появление таких количественных подходов к анализу отбора примечательно не только благодаря их технической применимости в изучении эволюции: они также являются признаком фундаментальных изменений в способах осмысления отбора биологами. Дарвиновская качественная идея, которая была выражена в абстрактной математической форме Фишером и впервые измерена с использованием генетических методов Добржанским и его учениками, теперь превратилась в прямо измеряемую статистическую характеристику ансамблей нуклеотидных сайтов. Такая трансформация концепции отбора сродни тому, как классическая термодинамика с ее абстрактными потоками превратилась в статистическую физику Больцмана и Гиббса (см. гл. 4).
Эгоистичные гены, мусорная ДНК и мобильные элементы
Эволюция путем дупликации генов и геномов: ортологи и паралоги
Прерывистое равновесие и несостоятельность градуализма
Пандативы, экзаптация, эволюция как ремесленник и ошибочность панглоссианской парадигмы эволюции
Эволюция в мире микробов и вирусов и трехдоменное древо жизни
Вирусы и рождение эволюционной геномики
• Некодирующие сайты, такие как интронные последовательности, могут использоваться как фон нейтральной эволюции при измерении отбора на синонимичных сайтах (Ks/Ki, где Ki – частота замен для интронных сайтов).
• Критерий Макдональда – Крейтмана (Aquadro, 1997; McDonald and Kreitman, 1991) широко используется для измерения отбора. Он сравнивает внутривидовые вариации (частота полиморфизма, P) с межвидовыми вариациями (дивергенция, D).
• Dn/Ds = Pn/Ps – нейтральная эволюция белковой последовательности.
• Dn/Ds < Pn/Ps – отсекающий отбор.
• Dn/Ds > Pn/Ps – положительный отбор.
Появление таких количественных подходов к анализу отбора примечательно не только благодаря их технической применимости в изучении эволюции: они также являются признаком фундаментальных изменений в способах осмысления отбора биологами. Дарвиновская качественная идея, которая была выражена в абстрактной математической форме Фишером и впервые измерена с использованием генетических методов Добржанским и его учениками, теперь превратилась в прямо измеряемую статистическую характеристику ансамблей нуклеотидных сайтов. Такая трансформация концепции отбора сродни тому, как классическая термодинамика с ее абстрактными потоками превратилась в статистическую физику Больцмана и Гиббса (см. гл. 4).
Эгоистичные гены, мусорная ДНК и мобильные элементы
Хоть это и редко утверждается без обиняков, классическая генетика предполагает, что почти все части генома (все нуклеотиды, если употреблять более современные, молекулярные термины) имеют определенные функции. Это неявно выраженное утверждение также важно и с точки зрения СТЭ, с ее панадаптационистским подходом. Однако это понимание подверглось сомнению еще в 1960-х и 1970-х годах по мере накопления данных об отсутствии прямой связи между размером генома и фенотипической сложностью организма. Даже с использованием приблизительных методов, доступных в то время, становилось ясно, что организмы с примерно одинаковым уровнем фенотипической сложности зачастую имеют геномы, на порядок различающиеся по размеру (так называемый парадокс гаплоидной величины). Этот парадокс был концептуально разрешен с помощью двух связанных друг с другом фундаментальных идей: эгоистичных генов и мусорной ДНК[21]. Концепция эгоистичных генов была предложена Ричардом Докинзом в одноименной книге, изданной в 1976 году (Dawkins, 2006). Резко отступив от организм-центричной парадигмы СТЭ, Докинз приходит к выводу, что естественный отбор может действовать не только на уровне организма в целом, но и на уровне индивидуального гена. Этот взгляд, поданный в умышленно провокационной манере, представляет геномы и организмы, по сути, средствами размножения генов.
Концепция эгоистичных генов породила множество важных выводов, и некоторые из них мы рассмотрим ниже в этой книге. Один из аспектов, имеющий непосредственное отношение к парадоксу гаплоидной величины, был всесторонне рассмотрен Фордом Дулиттлом и Кармен Сапиенцей (Doolittle and Sapienza, 1980), а также Лесли Оргелом и Фрэнсисом Криком (Orgel and Crick, 1980). Они предположили, что немалая или даже основная часть геномной ДНК (по крайней мере в сложных многоклеточных организмах) состоит из различных классов повторов, которые образуются в результате амплификации эгоистичных элементов – абсолютных паразитов, говоря хлестким языком Оргела и Крика. Другими словами, с точки зрения организма, большая часть геномной ДНК должна быть признана избыточной. Такой взгляд на геном в корне отличается от панселекционистской парадигмы, присущей СТЭ, в рамках которой большинство или даже все нуклеотиды в геноме подвержены влиянию отсекающего или положительного отбора, действующего на уровне организма.
Концептуально родственным важным открытием стало обнаружение транспозонов, или «прыгающих генов», сначала Барбарой Макклинток в 1940-х годах в растениях, а затем и в животных. Эти транспозоны стали затем известны как мобильные элементы (то есть генетические элементы, которые имеют тенденцию часто менять свое место в геноме; McClintock, 1984). Демонстрация вездесущности мобильных элементов привела к концепции высокодинамичных, постоянно меняющихся геномов задолго до рождения современной геномики[22].
Концепция эгоистичных генов породила множество важных выводов, и некоторые из них мы рассмотрим ниже в этой книге. Один из аспектов, имеющий непосредственное отношение к парадоксу гаплоидной величины, был всесторонне рассмотрен Фордом Дулиттлом и Кармен Сапиенцей (Doolittle and Sapienza, 1980), а также Лесли Оргелом и Фрэнсисом Криком (Orgel and Crick, 1980). Они предположили, что немалая или даже основная часть геномной ДНК (по крайней мере в сложных многоклеточных организмах) состоит из различных классов повторов, которые образуются в результате амплификации эгоистичных элементов – абсолютных паразитов, говоря хлестким языком Оргела и Крика. Другими словами, с точки зрения организма, большая часть геномной ДНК должна быть признана избыточной. Такой взгляд на геном в корне отличается от панселекционистской парадигмы, присущей СТЭ, в рамках которой большинство или даже все нуклеотиды в геноме подвержены влиянию отсекающего или положительного отбора, действующего на уровне организма.
Концептуально родственным важным открытием стало обнаружение транспозонов, или «прыгающих генов», сначала Барбарой Макклинток в 1940-х годах в растениях, а затем и в животных. Эти транспозоны стали затем известны как мобильные элементы (то есть генетические элементы, которые имеют тенденцию часто менять свое место в геноме; McClintock, 1984). Демонстрация вездесущности мобильных элементов привела к концепции высокодинамичных, постоянно меняющихся геномов задолго до рождения современной геномики[22].
Эволюция путем дупликации генов и геномов: ортологи и паралоги
СТЭ в полной мере унаследовала центральное положение дарвиновской теории, провозгласившей постепенные малые изменения единственно возможным материалом для эволюции. Однако эта концепция была поставлена под сомнение альтернативной концепцией эволюции дупликацией гена, разработанной Сусуму Оно в его классической книге 1970 года (Ohno, 1970). Мысль о том, что дупликация частей хромосом может служить одной из движущих сил эволюции, восходит к основателям современной количественной генетики, в частности к Фишеру и Холдейну[23]. Однако Оно первым предположил, что дупликация генов является основой эволюции геномов и организмов, и первым подвел качественную теорию под это положение. Начав с цитогенетических свидетельств полногеномной дупликации (ПГД) в начале эволюции хордовых, Оно выдвинул гипотезу о том, что дупликация генов является важным, если не единственным, путем эволюционии новых биологических функций. Согласно гипотезе Оно, дупликация гена высвобождает одну из копий от ограничений отсекающего отбора и, таким образом, эта копия получает потенциал развития новой функции (феномен, позднее названный неофункционализацией). Очевидно, что возникновение нового гена в результате дупликации, не говоря уже о дупликации геномного участка или ПГД, является огромным отличием от дарвиновских ничтожно малых изменений. Если такие крупные события в самом деле являются ключевыми для эволюции, то парадигма постепенных изменений в опасности. Позднейшие исследования дупликации генов, обсуждаемые далее в этой книге (см. гл. 8 и 9), привели к предположению о том, что неофункционализация вряд ли является основным путем эволюции дуплицированных генов. Однако факт остается фактом: дупликация, как важнейший механизм эволюции, бросает вызов градуализму.
Примерно в то же время, когда была издана книга Оно об эволюции путем дупликации генов, Уолтер Фитч опубликовал весьма плодотворную статью, всю значимость которой стало возможным оценить лишь в свете более поздних достижений геномики. Фитч исследовал понятие гомологии (общего предка) генов и провел различие между двумя классами гомологичных генов: ортологами и паралогами (Fitch, 1970). Ортологи – это гены, которые эволюционировали вертикально от одного предкового гена, принадлежащего общему предку сравниваемых организмов, тогда как паралоги – гены, эволюционировавшие в результате дупликации. Понятия ортологии и паралогии очевидным образом тесно связаны между собой и зависят от конкретной топологии филогенетического дерева данного семейства генов, так что дупликация в определенном узле дерева порождает новый набор паралогов в поддереве-потомке (подробнее см. в гл. 3). Более того, концептуальное определение паралогии осложнено специфичными для каждой линии эволюции потерей и горизонтальным переносом генов (см. гл. 5 и 7). Тем не менее, если не принимать во внимание эти осложнения, классификация гомологов Фитча остается центральной для эволюционной геномики[24].
Примерно в то же время, когда была издана книга Оно об эволюции путем дупликации генов, Уолтер Фитч опубликовал весьма плодотворную статью, всю значимость которой стало возможным оценить лишь в свете более поздних достижений геномики. Фитч исследовал понятие гомологии (общего предка) генов и провел различие между двумя классами гомологичных генов: ортологами и паралогами (Fitch, 1970). Ортологи – это гены, которые эволюционировали вертикально от одного предкового гена, принадлежащего общему предку сравниваемых организмов, тогда как паралоги – гены, эволюционировавшие в результате дупликации. Понятия ортологии и паралогии очевидным образом тесно связаны между собой и зависят от конкретной топологии филогенетического дерева данного семейства генов, так что дупликация в определенном узле дерева порождает новый набор паралогов в поддереве-потомке (подробнее см. в гл. 3). Более того, концептуальное определение паралогии осложнено специфичными для каждой линии эволюции потерей и горизонтальным переносом генов (см. гл. 5 и 7). Тем не менее, если не принимать во внимание эти осложнения, классификация гомологов Фитча остается центральной для эволюционной геномики[24].
Прерывистое равновесие и несостоятельность градуализма
Недостача межвидовых переходных форм в палеонтологической летописи – постоянная тема эволюционной биологии. Дарвин осознавал эту проблему и традиционно считал ее (так же как и палеонтологи, следующие дарвиновским традициям) следствием драматической неполноты этой летописи. Однако обширное накопление палеонтологических данных в XX веке мало помогло (если не сказать совершенно не помогло) в решении этой проблемы, что привело к возникновению иной точки зрения, сначала с появлением концепции квантовой эволюции Джорджа Гэйлорда Симпсона, затем оформившейся в концепцию прерывистого равновесия Стивена Джея Гулда и Нильса Элдриджа (Eldredge and Gould, 1997; Gould, 2002). Гулд и Элдридж собрали обширную доказательную базу, свидетельствующую о том, что история большинства видов животных, отраженная в палеонтологической летописи, соответствует в основном состоянию покоя – то есть, фактически, отсутствия изменений. Состояние покоя (стасис) перемежается «внезапным» исчезновением видов, последовательно замещаемых новыми. Следствием такой модели является очень быстрое в сравнении с продолжительностью стасиса видообразование; возникновение новых видов в определенной области является следствием миграции из области видообразования; градуалистское видообразование (постепенная трансформация видов в новые) – довольно редкий процесс. Такая модель прерывистого равновесия кажется применимой и к эволюции высших таксонов и зачастую обобщается до несостоятельности градуализма в целом, хотя правомерность такого вывода часто подвергается критике.
Пандативы, экзаптация, эволюция как ремесленник и ошибочность панглоссианской парадигмы эволюции
Пусть и неявно, но принципу градуализма был брошен вызов гипотезой Оно об эволюции генов и геномов путем дупликации, a затем, в явной форме, концепцией прерывистого равновесия. Адаптационистская программа эволюционной биологии подверглась решительной, сметающей все на своем пути атаке в статье 1979 года «Пандативы Святого Марка» Гулда и Левонтина (Gould and Lewontin, 1979), одной из самых необычных и влиятельных статей в истории биологии. Гулд и Левонтин саркастически описали адаптационистскую картину мира как панглоссианскую парадигму, названную так в честь примечательного персонажа вольтеровского «Кандида», который утверждал, что «все к лучшему в этом мире» [пер. Ф. Сологуба] (даже катастрофы). Гулд и Левонтин подчеркивали, что вместо того, чтобы стряпать на скорую руку «сказки просто так»[25] о правдоподобных адаптациях, эволюционным биологам следовало бы искать объяснение наблюдаемых черт организации биологических организмов исходя из плюралистского подхода, который принимает во внимание не только отбор, но также и внутренние ограничения, случайный дрейф и другие факторы. Метафора пандатива означает, что многие функционально важные элементы биологической организации вовсе не эволюционировали как специальные устройства для выполнения определенных функций, но скорее являются продуктами неадаптивных архитектурных ограничений, подобно пандативам (spandrels), появляющимся в арках соборов и других зданий исключительно вследствие требований конструкции, и могут использоваться для различных целей, например для украшения собора (см. рис. 2–2). Процессу использования пандативов в биологии было дано специальное название экзаптация, и Гулд провозгласил его важным путем эволюции (Gould, 1997a). Концепция пандативов связана с почти нейтральной теорией, но в каком-то смысле идет дальше и подходит ближе к сути эволюционного мышления, показывая, что даже те фенотипические черты, которые выглядят как типичные адаптации, не обязательно эволюционировали под давлением естественного отбора.
 Рис. 2–2. Один из пандативов базилики Святого Марка в Венеции. Фото Марии Шнитцмейер, Викисклад.
Рис. 2–2. Один из пандативов базилики Святого Марка в Венеции. Фото Марии Шнитцмейер, Викисклад.
В более ранней статье по сходной тематике Франсуа Жакоб (один из первооткрывателей регуляции генов и автор нескольких других плодотворных идей в бактериальной генетике, см. гл. 5) ввел метафору мастера-самоучки. Отталкиваясь прежде всего от сравнительного анализа механизмов развития, Жакоб положил в основу своих рассуждений, что эволюция действует не как инженер или дизайнер, а скорее как ремесленник-самоучка, причем чрезвычайно зависимый от предыдущего опыта при решении стоящих перед ним проблем: «Сложно проводить аналогии между естественным отбором и какими-то аспектами человеческого поведения. Однако если очень хочется поиграть в сравнения, то можно сказать, что естественный отбор работает не как изобретатель или инженер. Он работает как дилетант – мастер на все руки, который не знает точно, что он собирается создать, и при этом использует все, что подвернется под руку, будь то обрывки ниток, куски дерева или старые коробки; короче, он действует как тот мастеровой, который использует все, что есть в его распоряжении, чтобы сделать хоть что-то, лишь бы работало» (Jacob, 1977).
Ключевым выводом концепции ремесленника-самоучки становится то, что итоговый результат эволюции непредсказуем, или по крайней мере его невозможно предсказать, не зная в деталях всех предшествующих событий. Другими словами, если взять и «проиграть эволюционную пластинку заново» (любимая метафора Гулда) в некоем мысленном эксперименте, то результат будет отличен от того, что мы наблюдаем в реальности, возможно до неузнаваемости; мы вернемся к этому обсуждению позднее в этой книге (см. гл. 13).
В более ранней статье по сходной тематике Франсуа Жакоб (один из первооткрывателей регуляции генов и автор нескольких других плодотворных идей в бактериальной генетике, см. гл. 5) ввел метафору мастера-самоучки. Отталкиваясь прежде всего от сравнительного анализа механизмов развития, Жакоб положил в основу своих рассуждений, что эволюция действует не как инженер или дизайнер, а скорее как ремесленник-самоучка, причем чрезвычайно зависимый от предыдущего опыта при решении стоящих перед ним проблем: «Сложно проводить аналогии между естественным отбором и какими-то аспектами человеческого поведения. Однако если очень хочется поиграть в сравнения, то можно сказать, что естественный отбор работает не как изобретатель или инженер. Он работает как дилетант – мастер на все руки, который не знает точно, что он собирается создать, и при этом использует все, что подвернется под руку, будь то обрывки ниток, куски дерева или старые коробки; короче, он действует как тот мастеровой, который использует все, что есть в его распоряжении, чтобы сделать хоть что-то, лишь бы работало» (Jacob, 1977).
Ключевым выводом концепции ремесленника-самоучки становится то, что итоговый результат эволюции непредсказуем, или по крайней мере его невозможно предсказать, не зная в деталях всех предшествующих событий. Другими словами, если взять и «проиграть эволюционную пластинку заново» (любимая метафора Гулда) в некоем мысленном эксперименте, то результат будет отличен от того, что мы наблюдаем в реальности, возможно до неузнаваемости; мы вернемся к этому обсуждению позднее в этой книге (см. гл. 13).
Эволюция в мире микробов и вирусов и трехдоменное древо жизни
Вероятно, в ходе развития биологии наибольшее влияние на изменение представления об эволюции оказало распространение эволюционных исследований на мир микробов, а именно одноклеточных эукариот (протист), прокариот (бактерий и архей) и вирусов. Дарвиновское представление об эволюции и все достижения эволюционной биологии нескольких последующих десятилетий базировались исключительно на исследовании животных и растений, тогда как одноклеточные эукариоты (протисты) и бактерии (монеры) были сугубо номинально размещены у корня древа жизни Эрнстом Геккелем и его последователями. Хотя к 1950-м годам генетический анализ бактериофагов и бактерий продвинулся настолько, что стало очевидным, что эти формы жизни обладают эволюционирующими геномами, СТЭ не принимала во внимание эти открытия. То, что бактерии (не говоря уже о вирусах) эволюционируют по тому же самому принципу и с использованием тех же механизмов, что и животные и растения, отнюдь не очевидно, учитывая все их разительные биологические отличия от многоклеточных организмов, и в особенности из-за отсутствия у них типичного полового размножения и репродуктивной изоляции, ключевых для видообразования среди животных и растений.
Фактически прокариоты стали «видны» эволюционным биологам в 1977 году, после выхода революционной работы Вёзе и его коллег по филогенезу рРНК (Woese, 1987)[26]. Рассмотренное в общем контексте, открытие Вёзе является эпохально важным и, возможно, даже заслуживает сравнения с открытием структуры ДНК. Вёзе установил, что в одной молекулярной структуре, а именно последовательности нуклеотидов рРНК, выявляется очевидная консервативность во всем диапазоне клеточных форм жизни. Кроме того, чрезвычайно информативным оказался и филогенетический анализ этой универсальной консервативной молекулы: он показал, что рРНК, в некотором приближении, эволюционирует с постоянной скоростью, то есть подчиняется модели молекулярных часов. Это привело к еще одному важному открытию, ставшему одним из символов эволюционной биологии конца ХХ века, – трехдоменному древу жизни (см. рис. 2–3; Woese et al., 1990). Тремя доменами являются бактерии, археи и эукариоты. Домен архей был открыт Джорджем Фоксом и Вёзе сравнительным анализом рРНК, когда в новой группе ничем, казалось бы, не примечательных «бактерий» обнаружились существенные отличия как от остальных бактерий, так и от более сложных эукариотических организмов. В дополнение к разграничению трех доменов, Вёзе и его коллеги использовали филогенетический анализ рРНК для идентификации нескольких основных ветвей архей и бактерий (Woese, 1987). Из этого следовало, что эволюция прокариот столь же доступна для изучения, как эволюция сложных эукариот, – концепция, чуждая микробиологам до работы Вёзе (Stanier and Van Niel, 1962). Благодаря достижениям Вёзе, его сотрудников и последователей появилась все усиливающаяся тенденция приравнивать филогенетическое древо рРНК, с его трехдоменной структурой, к древу жизни Дарвина и Геккеля (Pace, 2009а, 2006). В течение нескольких лет после публикации открытий Вёзе стало ясно, что топологически древо рРНК (по крайней мере, в своих основных чертах) конгруэнтно деревьям некоторых из самых консервативных белков, таких как рибосомные белки, факторы трансляции, субъ единицы ДНК-зависимой РНК-полимеразы и мембранные АТФазы.
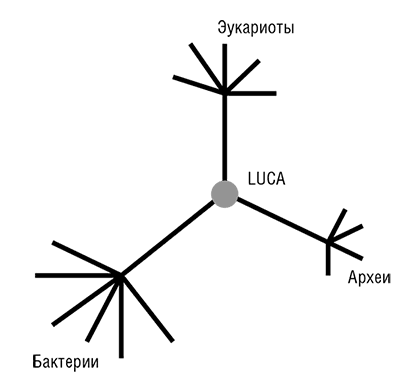 Рис. 2–3. Трехдоменное древо жизни Вёзе.
Рис. 2–3. Трехдоменное древо жизни Вёзе.
Две группы исследователей независимо друг от друга пришли к блестящей идее о том, как определить положение корня в эволюционном дереве, которое до этого было бескорневым (рис. 2–3). Для этой цели можно использовать древние паралоги, которые представлены в (почти) всех организмах и, таким образом, можно с уверенностью заключить, возникли в результате дупликации, предшествующей последнему общему предку всех живых организмов (LUCA). Когда дерево строится совместно для двух паралогичных множеств древних ортологов, положение корня между ними определено однозначно, и таким образом корень может быть выведен для каждого из множеств ортологов (см. рис. 2–4; Gogarten et al., 1989; Iwabe et al., 1989). Результаты анализа двух пар древних паралогов, факторов трансляции и субъединиц мембранных АТФаз были полностью совместимы и поместили корень на бактериальную ветвь, установив таким образом кладу архей-эукариотов (см. рис. 2–4). Тем не менее даже в догеномную эпоху было ясно, что не все деревья белок-кодирующих генов имеют ту же топологию, что и дерево рРНК; причины этих отличий оставались неясными и, как предполагалось, включали (за исключением артефактов метода) горизонтальный перенос генов (ГПГ. Smith et al., 1992). Эти расхождения оставались лишь интересным дополнением к трехдоменному ДЖ, но все резко изменилось с наступлением эры геномики.
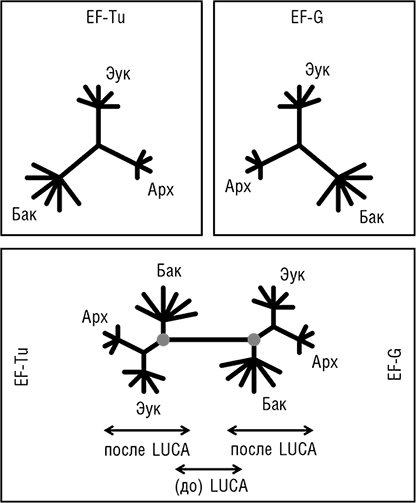 Рис. 2–4. Определение положения корня в трехдоменном древе жизни с помощью древних паралогов. Схематически показаны филогенетические деревья двух широко распространенных факторов инициации трансляции EF-Tu и EF-G, реконструированные независимо (верхние диаграммы) и совместно (нижняя диаграмма). Кружками обозначено вычисленное положение корня в каждом из двух деревьев.
Рис. 2–4. Определение положения корня в трехдоменном древе жизни с помощью древних паралогов. Схематически показаны филогенетические деревья двух широко распространенных факторов инициации трансляции EF-Tu и EF-G, реконструированные независимо (верхние диаграммы) и совместно (нижняя диаграмма). Кружками обозначено вычисленное положение корня в каждом из двух деревьев.
Фактически прокариоты стали «видны» эволюционным биологам в 1977 году, после выхода революционной работы Вёзе и его коллег по филогенезу рРНК (Woese, 1987)[26]. Рассмотренное в общем контексте, открытие Вёзе является эпохально важным и, возможно, даже заслуживает сравнения с открытием структуры ДНК. Вёзе установил, что в одной молекулярной структуре, а именно последовательности нуклеотидов рРНК, выявляется очевидная консервативность во всем диапазоне клеточных форм жизни. Кроме того, чрезвычайно информативным оказался и филогенетический анализ этой универсальной консервативной молекулы: он показал, что рРНК, в некотором приближении, эволюционирует с постоянной скоростью, то есть подчиняется модели молекулярных часов. Это привело к еще одному важному открытию, ставшему одним из символов эволюционной биологии конца ХХ века, – трехдоменному древу жизни (см. рис. 2–3; Woese et al., 1990). Тремя доменами являются бактерии, археи и эукариоты. Домен архей был открыт Джорджем Фоксом и Вёзе сравнительным анализом рРНК, когда в новой группе ничем, казалось бы, не примечательных «бактерий» обнаружились существенные отличия как от остальных бактерий, так и от более сложных эукариотических организмов. В дополнение к разграничению трех доменов, Вёзе и его коллеги использовали филогенетический анализ рРНК для идентификации нескольких основных ветвей архей и бактерий (Woese, 1987). Из этого следовало, что эволюция прокариот столь же доступна для изучения, как эволюция сложных эукариот, – концепция, чуждая микробиологам до работы Вёзе (Stanier and Van Niel, 1962). Благодаря достижениям Вёзе, его сотрудников и последователей появилась все усиливающаяся тенденция приравнивать филогенетическое древо рРНК, с его трехдоменной структурой, к древу жизни Дарвина и Геккеля (Pace, 2009а, 2006). В течение нескольких лет после публикации открытий Вёзе стало ясно, что топологически древо рРНК (по крайней мере, в своих основных чертах) конгруэнтно деревьям некоторых из самых консервативных белков, таких как рибосомные белки, факторы трансляции, субъ единицы ДНК-зависимой РНК-полимеразы и мембранные АТФазы.
Две группы исследователей независимо друг от друга пришли к блестящей идее о том, как определить положение корня в эволюционном дереве, которое до этого было бескорневым (рис. 2–3). Для этой цели можно использовать древние паралоги, которые представлены в (почти) всех организмах и, таким образом, можно с уверенностью заключить, возникли в результате дупликации, предшествующей последнему общему предку всех живых организмов (LUCA). Когда дерево строится совместно для двух паралогичных множеств древних ортологов, положение корня между ними определено однозначно, и таким образом корень может быть выведен для каждого из множеств ортологов (см. рис. 2–4; Gogarten et al., 1989; Iwabe et al., 1989). Результаты анализа двух пар древних паралогов, факторов трансляции и субъединиц мембранных АТФаз были полностью совместимы и поместили корень на бактериальную ветвь, установив таким образом кладу архей-эукариотов (см. рис. 2–4). Тем не менее даже в догеномную эпоху было ясно, что не все деревья белок-кодирующих генов имеют ту же топологию, что и дерево рРНК; причины этих отличий оставались неясными и, как предполагалось, включали (за исключением артефактов метода) горизонтальный перенос генов (ГПГ. Smith et al., 1992). Эти расхождения оставались лишь интересным дополнением к трехдоменному ДЖ, но все резко изменилось с наступлением эры геномики.
Вирусы и рождение эволюционной геномики
Эволюционная геномика родилась более чем за десять лет до исторического заявления о секвенировании первого бактериального генома. С меньшей помпой (но и не в безвестности) было секвенировано несколько небольших (в рамках 4—100 Кб) различных вирусных геномов, и были разработаны принципы сравнения геномов, наряду с практическими вычислительными методами. Вирусы являются облигатными внутриклеточными паразитами, и вирусные геномы намного меньше и качественно отличаются от геномов клеточных форм жизни. Вирусы обычно не лишены некоторых классов генов, вездесущих и незаменимых в клеточных организмах, таких как гены компонентов системы трансляции и биогенеза мембран. Тем не менее вирусы следуют своей собственной «биологической стратегии» и кодируют все субъединицы вириона, а также по крайней мере некоторые белки, участвующие в репликации вирусного генома. (Одна из центральных тем этой книги – ключевая роль вирусов в эволюции биосферы, так что я возвращаюсь к ней достаточно подробно в гл. 10[27].) Несмотря на быструю эволюцию последовательности генома, характерную для вирусов (в особенности вирусов с РНК-геномом), эти ранние сравнительные геномные исследования успешно выявили множества генов, консервативных в больших группах вирусов (Koonin and Dolja, 1993). Возможность структурного и функционального картирования всего генома определенной формы жизни была реализована в этих исследованиях впервые, и это стало краеугольным камнем эволюционной геномики. Кроме того, было сделано непредвиденное и важное обобщение: в то время как некоторые гены консервативны для удивительно обширного разнообразия вирусов, архитектура генома, структура вириона и биологические свойства вирусов демонстрируют гораздо большую пластичность (см. гл. 5 и 10, где этот вопрос обсуждается подробнее).