Страница:
У млекопитающих половое запечатление изучено главным образом у копытных и грызунов. Имеются, например, указания на то, что скрещивание между ламами викуньей и альпакой возможно лишь в том случае, если новорожденному представителю первого вида дают в качестве кормилицы самку альпака. Немецкие этологи Д. и Х. Мюллер-Шварце показали, что у чернохвостого оленя в половом запечатлении главную роль играют запахи. Группой итальянских исследователей проводились опыты, в которых мыши-родители опрыскивались пахучими веществами. В результате их детеныши, будучи половозрелыми, оказались не в состоянии различать пол сверстников, т.е. не могли найти полового партнера. Аналогичные результаты были получены и у крыс. Опыты с применением «приемных родителей» производились и с морскими свинками. Эффект полового запечатления на чужой вид был выявлен у самцов, отделенных в течение первой недели жизни от своих матерей.
Интересно отметить, что половое запечатление может иметь место и у взрослых особей. Доказано это для некоторых рыб, например меченосца (Xiphohorus maculatus) - известной аквариумной рыбки из отряда карпозубых. Самцы этого вида, как показали шведские этологи А. Ферне и С.Сьеландер, предпочитают самок той окраски, которую им приходилось видеть в течение двух месяцев после наступления половозрелости, но не раньше, как это имеет место в описанных выше случаях полового запечатления.
Подводя итог обзору процессов запечатления, важно подчеркнуть, что общим и наиболее существенным для всех форм запечатления является быстрая постнатальная достройка врожденного поведения, точнее, врожденных пусковых механизмов путем их дополнения индивидуально приобретаемыми компонентами. В результате инстинктивное поведение конкретизируется на определенных, индивидуально опознаваемых объектах, чем и обеспечивается эффективность выполнения инстинктивных действий.
Раннее факультативное научение
Как самостоятельная категория приобретения индивидуального опыта факультативное научение играет в раннем онтогенезе еще значительно меньшую роль, чем на последующих этапах онтогенеза. Это объясняется тем, что оно первоначально лишь дополняет облигатное научение.
Экспериментально факультативный компонент раннего научения можно выявить, воздействуя на детенышей искусственными стимулами, которые или совершенно не встречаются в нормальной среде обитания данного вида, или хотя и могут встречаться, не обладают сколько-нибудь существенной биологической валентностью и не обусловливают определенную инстинктивную реакцию. Кроме того, этот компонент может быть выявлен путем обучения животных действиям, не представленным в их видотипичном поведении.
В качестве примера можно привести эксперимент, в котором крысят в возрасте 5, 7, 9 и 11 дней обучали с помощью электрического раздражения преодолевать коридор, причем ток выключался только при достижении крысятами стенки в конце коридора. При всех прочих оборонительных реакциях (поворотах, попытках забраться на стенку, опрокидывании на бок) действие болевого раздражителя продолжалось. У всех подопытных животных по мере тренировки число таких реакций уменьшалось, но только 7- 11 -дневные крысята научились все быстрее находить правильное направление к стенке в конце коридора и тем самым скорее избавляться от действия тока. Это говорит о том, что до 5-дневного возраста факультативное научение проявляется в еще весьма ограниченных рамках и неспособно помочь животному в выборе адекватного направления действия. Еще позже у крысят появляются факультативные реакции на положительные раздражители. Так, например, приблизительно только с 20-дневного возраста их можно научить нажимать на рычаг за пищевое вознаграждение. Однако, по данным ряда исследователей, оптимальный период раннего постнатального научения начинается у них в месячном возрасте.
У хищных млекопитающих отмечаются аналогичные сроки формирования первых положительных навыков. У щенков было установлено, что прогрессивное развитие способности к формированию навыков в раннем постнатальном онтогенезе непосредственно зависит от развития кратковременной памяти, что в свою очередь связано с развитием коркового торможения (Р.Г.Лозовская и Г.А.Образцова).
У других незрелорождающихся млекопитающих, которые появляются, однако, на свет с уже вполне функционирующими дистантными рецепторами, факультативное научение начинается значительно раньше. Так, по данным советской исследовательницы Н.А.Тих, много лет изучающей психику обезьян в сравнительно-психологическом аспекте, у низших обезьян в условиях искусственного выращивания уже на 3-4-й день после рождения появляются специфические реакции на вид рожка, из которого кормят детеныша, в частности своеобразные движения рук для его обхватывания (после 40-50 кормлений). Экспериментальный условный рефлекс на звук Тих удалось выработать у детеныша гамадрила в возрасте 29 дней.
Необходимо, конечно, учесть, что первые условные рефлексы на тактильные, термические или проприоцептивные раздражители образуются и у других млекопитающих в первые же дни после их рождения. Не исключено, следовательно, что в сфере действия таких раздражителей с самого начала постнатального развития может иметь место не только облигатное, но и факультативное научение даже у животных, рождающихся в крайне незрелом состоянии. Само собой разумеется, что именно так обстоит дело у всех зрелорождающихся животных.
Что касается низших позвоночных, то, как показали некоторые исследования, в частности В.А.Трошихина, экспериментальная выработка стойких условных рефлексов на искусственные раздражители удается уже у мальков рыб. Так, для образования условного пищевого двигательного рефлекса на вспыхивание электрической лампочки у малька карпа достаточно в среднем 4-6 сочетаний, причем положительная условная реакция остается стойкой в течение 100 повторений светового раздражителя без пищевого подкрепления.
Взаимоотношения между компонентами раннего постнатального развития поведения
Раннее формирование общения
Интересно отметить, что половое запечатление может иметь место и у взрослых особей. Доказано это для некоторых рыб, например меченосца (Xiphohorus maculatus) - известной аквариумной рыбки из отряда карпозубых. Самцы этого вида, как показали шведские этологи А. Ферне и С.Сьеландер, предпочитают самок той окраски, которую им приходилось видеть в течение двух месяцев после наступления половозрелости, но не раньше, как это имеет место в описанных выше случаях полового запечатления.
Подводя итог обзору процессов запечатления, важно подчеркнуть, что общим и наиболее существенным для всех форм запечатления является быстрая постнатальная достройка врожденного поведения, точнее, врожденных пусковых механизмов путем их дополнения индивидуально приобретаемыми компонентами. В результате инстинктивное поведение конкретизируется на определенных, индивидуально опознаваемых объектах, чем и обеспечивается эффективность выполнения инстинктивных действий.
Раннее факультативное научение
Как самостоятельная категория приобретения индивидуального опыта факультативное научение играет в раннем онтогенезе еще значительно меньшую роль, чем на последующих этапах онтогенеза. Это объясняется тем, что оно первоначально лишь дополняет облигатное научение.
Экспериментально факультативный компонент раннего научения можно выявить, воздействуя на детенышей искусственными стимулами, которые или совершенно не встречаются в нормальной среде обитания данного вида, или хотя и могут встречаться, не обладают сколько-нибудь существенной биологической валентностью и не обусловливают определенную инстинктивную реакцию. Кроме того, этот компонент может быть выявлен путем обучения животных действиям, не представленным в их видотипичном поведении.
В качестве примера можно привести эксперимент, в котором крысят в возрасте 5, 7, 9 и 11 дней обучали с помощью электрического раздражения преодолевать коридор, причем ток выключался только при достижении крысятами стенки в конце коридора. При всех прочих оборонительных реакциях (поворотах, попытках забраться на стенку, опрокидывании на бок) действие болевого раздражителя продолжалось. У всех подопытных животных по мере тренировки число таких реакций уменьшалось, но только 7- 11 -дневные крысята научились все быстрее находить правильное направление к стенке в конце коридора и тем самым скорее избавляться от действия тока. Это говорит о том, что до 5-дневного возраста факультативное научение проявляется в еще весьма ограниченных рамках и неспособно помочь животному в выборе адекватного направления действия. Еще позже у крысят появляются факультативные реакции на положительные раздражители. Так, например, приблизительно только с 20-дневного возраста их можно научить нажимать на рычаг за пищевое вознаграждение. Однако, по данным ряда исследователей, оптимальный период раннего постнатального научения начинается у них в месячном возрасте.
У хищных млекопитающих отмечаются аналогичные сроки формирования первых положительных навыков. У щенков было установлено, что прогрессивное развитие способности к формированию навыков в раннем постнатальном онтогенезе непосредственно зависит от развития кратковременной памяти, что в свою очередь связано с развитием коркового торможения (Р.Г.Лозовская и Г.А.Образцова).
У других незрелорождающихся млекопитающих, которые появляются, однако, на свет с уже вполне функционирующими дистантными рецепторами, факультативное научение начинается значительно раньше. Так, по данным советской исследовательницы Н.А.Тих, много лет изучающей психику обезьян в сравнительно-психологическом аспекте, у низших обезьян в условиях искусственного выращивания уже на 3-4-й день после рождения появляются специфические реакции на вид рожка, из которого кормят детеныша, в частности своеобразные движения рук для его обхватывания (после 40-50 кормлений). Экспериментальный условный рефлекс на звук Тих удалось выработать у детеныша гамадрила в возрасте 29 дней.
Необходимо, конечно, учесть, что первые условные рефлексы на тактильные, термические или проприоцептивные раздражители образуются и у других млекопитающих в первые же дни после их рождения. Не исключено, следовательно, что в сфере действия таких раздражителей с самого начала постнатального развития может иметь место не только облигатное, но и факультативное научение даже у животных, рождающихся в крайне незрелом состоянии. Само собой разумеется, что именно так обстоит дело у всех зрелорождающихся животных.
Что касается низших позвоночных, то, как показали некоторые исследования, в частности В.А.Трошихина, экспериментальная выработка стойких условных рефлексов на искусственные раздражители удается уже у мальков рыб. Так, для образования условного пищевого двигательного рефлекса на вспыхивание электрической лампочки у малька карпа достаточно в среднем 4-6 сочетаний, причем положительная условная реакция остается стойкой в течение 100 повторений светового раздражителя без пищевого подкрепления.
Взаимоотношения между компонентами раннего постнатального развития поведения
Все компоненты раннего постнатального развития поведения переплетаются друг с другом, в результате чего это развитие приобретает очень сложный характер. Проанализируем эти связи и взаимозависимости на приведенном выше примере поведения вылупившихся гоголят.
В поведении этих птенцов прежде всего обращает на себя внимание врожденная (инстинктивная) двигательная реакция - подскакивание птенцов к выходу из дупла в ответ на призывные крики матери, а также врожденное узнавание светлого отверстия как цели прыжка. Врожденный характер подскакивания подтверждается работами норвежского исследователя Т.Бьерке, установившего, что для его появления необходим определенный уровень физиологической «зрелости» птенца. Но едва ли имеет здесь место также всецело врожденное узнавание призывного крика, так как еще до вылупления птенцов между ними и материнской особью, вероятно, устанавливается уже знакомый нам взаимный акустический контакт, как это доказано для других зрелорождающихся птиц. Следовательно, нельзя исключить возможность своего рода акустически-кинестетического эмбрионального запечатления голоса матери, т.е. пренатального облигатного научения, как его теперь можно назвать. С другой стороны, совершенно очевидно, что птенцы не могли в период эмбриогенеза тренироваться в целенаправленных прыжках.
Далее существенно, что гоголята реагируют на звуки матери еще до того, как увидят ее: только высунувшись из дупла, они впервые видят ее летящей перед собой. Также врожденная реакция спрыгивания на землю непосредственно не связана с ее присутствием (хотя, возможно, и стимулируется ее видом и голосом), так как подобным же образом ведут себя и гоголята, выращенные в инкубаторе, когда они оказываются на светлом месте (наблюдения Фабри).
Завершается этот первый постэмбриональный этап собственно реакцией следования утят за удаляющейся к водоему матерью (и друг за другом). Эта реакция наступает в ответ на вид двигающейся (в данном случае ходящей) птицы в сочетании с издаваемыми ею звуками, что четко доказано экспериментами, проведенными многочисленными исследователями не только на утятах, но и на цыплятах и птенцах других зрелорождающихся птиц.
Выше уже отмечалось, что конкретное значение запечатления типа реакции следования состоит в индивидуальном узнавании особенно важного компонента среды - родительской особи. Ясно, что это не может осуществляться только врожденным образом, так как распознаваться и запоминаться должны в данном случае не видовые, а именно индивидуальные признаки, т.е. частные отклонения от видотипичной нормы. Объектами врожденного узнавания могут быть лишь общие всем представителям данного вида признаки, а между тем детеныш должен быть в состоянии отличить своего родителя как раз от других особей того же вида. Этому можно только научиться, на это и направлено облигатное научение.
В некоторых случаях, правда, видотипичные признаки, хотя бы частично, тоже фиксируются в памяти детеныша путем запечатления. Особенно, как мы видели, это проявляется при половом запечатлении. В этом состоит одно из важнейших отличий полового запечатления.
Важно отметить, что процессы раннего облигатного научения, прежде всего запечатления, всегда сочетаются с врожденным узнаванием. При этом врожденное узнавание направлено на общие существенные признаки объектов видотипичных действий, путем же облигатного научения эти действия конкретизируются на частном объекте. Так, при реакции следования восприятие детенышем самой подвижности родительской особи (как и любого другого впервые увиденного объекта) является врожденным. Это обусловлено тем, что подвижность представляет собой в данном случае существенный признак объекта запечатления. Индивидуальные же отличительные признаки родительской особи (т.е. объекта видотипичного действия) заучиваются путем запечатления. Следовательно, слияние врожденного поведения (врожденного узнавания) с облигатным научением служит основой дальнейшего развития видотипичного поведения.
Вместе с тем в запечатлении содержатся элементы и факультативного научения. Так, например, птенцы одного и того же выводка могут научиться узнавать родительскую особь по разным ее признакам, и при этом не имеет значения, по каким конкретно признакам ориентируется тот или иной птенец. Облигатным, обязательным здесь является то, что каждый птенец должен вообще научиться узнавать родителя по каким-либо его индивидуальным признакам.
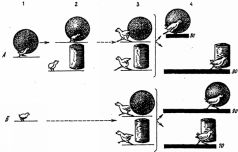
Сочетание врожденных и приобретаемых компонентов сказывается здесь еще и в другом. Как показали эксперименты, проведенные Фабри на цыплятах, первоначальный эффект врожденного узнавания при реакции следования недолговечен и без постоянного упражнения довольно быстро угасает. Однако этот эффект, т.е. следование, можно легко восстановить путем факультативного научения, направленного на объект запечатления. В упомянутых экспериментах объектом запечатления служил для одних цыплят синий шар, для других - желтый цилиндр. После первых же предъявлений цыплята стали следовать за этими объектами не менее интенсивно и продолжительно, чем в обычных условиях за курицей. Но даже наиболее активные подопытные цыплята переставали следовать за объектом запечатления уже через несколько дней, если прекращались ежедневные тренировки. В обычных же условиях, как известно, эта реакция сохраняется в течение ряда недель. Однако и в искусственных условиях эксперимента цыплята тем дольше продолжали следовать за объектом запечатления, чем больше было всего пробежек, т.е. длительность сохранения реакции следования оказалась прямо пропорциональной интенсивности локомоторной активности (частоте пробежек).
В дальнейшем, уже после того, как реакция следования у цыплят совсем исчезла, у них в специальной установке были выработаны навыки следования за бывшим объектом следования - синим шаром или соответственно желтым цилиндром. Следование за этим объектом подкреплялось пищей. Другими словами, следование было заново выработано у цыплят, но путем факультативного научения.
Результаты этой части эксперимента показали, что у цыплят, у которых раньше была сформирована реакция следования путем запечатления, этот навык образовался намного быстрее, чем у цыплят, которые до этого вообще не подвергались запечатлению, т.е. не имели возможности следовать за каким-либо подвижным объектом или же следовали за объектом запечатления другой группы подопытных животных (для цыплят, запечатленных на синий шар, это был желтый цилиндр и наоборот, - рис. 21).
Из этих данных, казалось бы, следует, что врожденное действие (само следование), протекание которого не зависит от побочных внешних факторов и сроки проявления которого генетически фиксированы, угасает без упражнения так же, как это происходит с навыками, а с другой стороны, навык, факультативное научение, может вновь «оживить» эту инстинктивную реакцию. Объясняется это, очевидно, тем, что существует не одна, а две генетически различные, но функционально тождественные формы следования - истинно врожденная и индивидуально приобретаемая, причем вторая форма образуется путем чисто факультативного научения (ведь курица подкрепляет следование своих цыплят кормом и обогревом). Очевидно также, что первая форма со временем все больше перерастает во вторую, иными словами, реакция следования претерпевает качественную перестройку, теряя свою первоначальную врожденную сущность. Одновременно врожденное узнавание переходит в благоприобретаемое узнавание, основанное на индивидуальном опыте.
Рис. 21. Формирование следования за подвижным объектом путем запечатления и факультативного научения (опыты Фабри). А -экспериментальная группа цыплят, Б - контрольная группа цыплят. 1- выработка и тренировка реакции следования за синим шаром путем запечатления (1-4-е сутки после вылупления);
2 - первый контрольный опыт; цыплята следуют за объектом запечатления (синим шаром), но не за другим подвижным объектом (желтым цилиндром) (5-е сутки); 3 - второй контрольный опыт (26-е сутки), проведенный после трехнедельного перерыва, во время которого цыплята находились в обычных условиях, но не имели возможности видеть шар и цилиндр. В результате цыплята не только не следуют за этими движущимися объектами, но даже пугаются их. Так же реагируют на движущиеся шар и цилиндр контрольные (не подвергшиеся запечатлению) цыплята; 4 - выработка навыка следования за двигающимися шаром или желтым цилиндром при одновременном их предъявлении (27-31-е сутки) путем факультативного научения. Столбики с цифрами показывают среднее число сочетаний, необходимых для формирования устойчивого следования
Все это указывает на значение раннего факультативного (а не только облигатного) научения для полноценного проявления инстинктивных элементов поведения. Иными словами, уже на первых этапах постнатального развития врожденное поведение нуждается в участии научения в обеих его формах, которые при этом сливаются с ним. Этим прежде всего и характеризуется сущность всего поведения животных на ранних этапах постнатального онтогенеза.
В поведении этих птенцов прежде всего обращает на себя внимание врожденная (инстинктивная) двигательная реакция - подскакивание птенцов к выходу из дупла в ответ на призывные крики матери, а также врожденное узнавание светлого отверстия как цели прыжка. Врожденный характер подскакивания подтверждается работами норвежского исследователя Т.Бьерке, установившего, что для его появления необходим определенный уровень физиологической «зрелости» птенца. Но едва ли имеет здесь место также всецело врожденное узнавание призывного крика, так как еще до вылупления птенцов между ними и материнской особью, вероятно, устанавливается уже знакомый нам взаимный акустический контакт, как это доказано для других зрелорождающихся птиц. Следовательно, нельзя исключить возможность своего рода акустически-кинестетического эмбрионального запечатления голоса матери, т.е. пренатального облигатного научения, как его теперь можно назвать. С другой стороны, совершенно очевидно, что птенцы не могли в период эмбриогенеза тренироваться в целенаправленных прыжках.
Далее существенно, что гоголята реагируют на звуки матери еще до того, как увидят ее: только высунувшись из дупла, они впервые видят ее летящей перед собой. Также врожденная реакция спрыгивания на землю непосредственно не связана с ее присутствием (хотя, возможно, и стимулируется ее видом и голосом), так как подобным же образом ведут себя и гоголята, выращенные в инкубаторе, когда они оказываются на светлом месте (наблюдения Фабри).
Завершается этот первый постэмбриональный этап собственно реакцией следования утят за удаляющейся к водоему матерью (и друг за другом). Эта реакция наступает в ответ на вид двигающейся (в данном случае ходящей) птицы в сочетании с издаваемыми ею звуками, что четко доказано экспериментами, проведенными многочисленными исследователями не только на утятах, но и на цыплятах и птенцах других зрелорождающихся птиц.
Выше уже отмечалось, что конкретное значение запечатления типа реакции следования состоит в индивидуальном узнавании особенно важного компонента среды - родительской особи. Ясно, что это не может осуществляться только врожденным образом, так как распознаваться и запоминаться должны в данном случае не видовые, а именно индивидуальные признаки, т.е. частные отклонения от видотипичной нормы. Объектами врожденного узнавания могут быть лишь общие всем представителям данного вида признаки, а между тем детеныш должен быть в состоянии отличить своего родителя как раз от других особей того же вида. Этому можно только научиться, на это и направлено облигатное научение.
В некоторых случаях, правда, видотипичные признаки, хотя бы частично, тоже фиксируются в памяти детеныша путем запечатления. Особенно, как мы видели, это проявляется при половом запечатлении. В этом состоит одно из важнейших отличий полового запечатления.
Важно отметить, что процессы раннего облигатного научения, прежде всего запечатления, всегда сочетаются с врожденным узнаванием. При этом врожденное узнавание направлено на общие существенные признаки объектов видотипичных действий, путем же облигатного научения эти действия конкретизируются на частном объекте. Так, при реакции следования восприятие детенышем самой подвижности родительской особи (как и любого другого впервые увиденного объекта) является врожденным. Это обусловлено тем, что подвижность представляет собой в данном случае существенный признак объекта запечатления. Индивидуальные же отличительные признаки родительской особи (т.е. объекта видотипичного действия) заучиваются путем запечатления. Следовательно, слияние врожденного поведения (врожденного узнавания) с облигатным научением служит основой дальнейшего развития видотипичного поведения.
Вместе с тем в запечатлении содержатся элементы и факультативного научения. Так, например, птенцы одного и того же выводка могут научиться узнавать родительскую особь по разным ее признакам, и при этом не имеет значения, по каким конкретно признакам ориентируется тот или иной птенец. Облигатным, обязательным здесь является то, что каждый птенец должен вообще научиться узнавать родителя по каким-либо его индивидуальным признакам.
Сочетание врожденных и приобретаемых компонентов сказывается здесь еще и в другом. Как показали эксперименты, проведенные Фабри на цыплятах, первоначальный эффект врожденного узнавания при реакции следования недолговечен и без постоянного упражнения довольно быстро угасает. Однако этот эффект, т.е. следование, можно легко восстановить путем факультативного научения, направленного на объект запечатления. В упомянутых экспериментах объектом запечатления служил для одних цыплят синий шар, для других - желтый цилиндр. После первых же предъявлений цыплята стали следовать за этими объектами не менее интенсивно и продолжительно, чем в обычных условиях за курицей. Но даже наиболее активные подопытные цыплята переставали следовать за объектом запечатления уже через несколько дней, если прекращались ежедневные тренировки. В обычных же условиях, как известно, эта реакция сохраняется в течение ряда недель. Однако и в искусственных условиях эксперимента цыплята тем дольше продолжали следовать за объектом запечатления, чем больше было всего пробежек, т.е. длительность сохранения реакции следования оказалась прямо пропорциональной интенсивности локомоторной активности (частоте пробежек).
В дальнейшем, уже после того, как реакция следования у цыплят совсем исчезла, у них в специальной установке были выработаны навыки следования за бывшим объектом следования - синим шаром или соответственно желтым цилиндром. Следование за этим объектом подкреплялось пищей. Другими словами, следование было заново выработано у цыплят, но путем факультативного научения.
Результаты этой части эксперимента показали, что у цыплят, у которых раньше была сформирована реакция следования путем запечатления, этот навык образовался намного быстрее, чем у цыплят, которые до этого вообще не подвергались запечатлению, т.е. не имели возможности следовать за каким-либо подвижным объектом или же следовали за объектом запечатления другой группы подопытных животных (для цыплят, запечатленных на синий шар, это был желтый цилиндр и наоборот, - рис. 21).
Из этих данных, казалось бы, следует, что врожденное действие (само следование), протекание которого не зависит от побочных внешних факторов и сроки проявления которого генетически фиксированы, угасает без упражнения так же, как это происходит с навыками, а с другой стороны, навык, факультативное научение, может вновь «оживить» эту инстинктивную реакцию. Объясняется это, очевидно, тем, что существует не одна, а две генетически различные, но функционально тождественные формы следования - истинно врожденная и индивидуально приобретаемая, причем вторая форма образуется путем чисто факультативного научения (ведь курица подкрепляет следование своих цыплят кормом и обогревом). Очевидно также, что первая форма со временем все больше перерастает во вторую, иными словами, реакция следования претерпевает качественную перестройку, теряя свою первоначальную врожденную сущность. Одновременно врожденное узнавание переходит в благоприобретаемое узнавание, основанное на индивидуальном опыте.
Рис. 21. Формирование следования за подвижным объектом путем запечатления и факультативного научения (опыты Фабри). А -экспериментальная группа цыплят, Б - контрольная группа цыплят. 1- выработка и тренировка реакции следования за синим шаром путем запечатления (1-4-е сутки после вылупления);
2 - первый контрольный опыт; цыплята следуют за объектом запечатления (синим шаром), но не за другим подвижным объектом (желтым цилиндром) (5-е сутки); 3 - второй контрольный опыт (26-е сутки), проведенный после трехнедельного перерыва, во время которого цыплята находились в обычных условиях, но не имели возможности видеть шар и цилиндр. В результате цыплята не только не следуют за этими движущимися объектами, но даже пугаются их. Так же реагируют на движущиеся шар и цилиндр контрольные (не подвергшиеся запечатлению) цыплята; 4 - выработка навыка следования за двигающимися шаром или желтым цилиндром при одновременном их предъявлении (27-31-е сутки) путем факультативного научения. Столбики с цифрами показывают среднее число сочетаний, необходимых для формирования устойчивого следования
Все это указывает на значение раннего факультативного (а не только облигатного) научения для полноценного проявления инстинктивных элементов поведения. Иными словами, уже на первых этапах постнатального развития врожденное поведение нуждается в участии научения в обеих его формах, которые при этом сливаются с ним. Этим прежде всего и характеризуется сущность всего поведения животных на ранних этапах постнатального онтогенеза.
Раннее формирование общения
Раннее формирование общения у птиц
Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда, коммуникативные компоненты поведения уже неоднократно встречались нами при ознакомлении с разными аспектами онтогенеза поведения, особенно в процессах запечатления, одним из определяющих факторов которого и является общение.
У птиц, как было уже показано (в частности, в предыдущей главе на кайрах), установление акустических контактов между родительской особью и ее потомством начинается уже на эмбриональной стадии развития птенцов путем пренатального облигатного научения (эмбрионального запечатления). У кайр, например, этот процесс происходит в форме установления акустико-кинестетических связей. В результате этого запечатления птенец подбегает к источнику запечатленного звука. Эта реакция обнаруживается даже у только что вылупившегося птенца: услышав запечатленный звук, он подбегает к родительской особи (в эксперименте к замещающему ее громкоговорителю), издает ответный крик и залезает под нее (или под аппарат). Если же из громкоговорителя после этого раздается другой звук (чужой зов), то птенец немедленно выскакивает из-под него и прячется в укрытии (опыты Чанц). Таким образом, коммуникативный компонент врожденного пускового механизма достраивается в данном случае в ходе эмбрионального запечатления.
Вместе стем и у родительской особи происходит запечатление индивидуальных особенностей голоса еще не вылупившегося птенца, что обеспечивает индивидуальное опознавание последнего уже к моменту вылупления. Подобное «обратное запечатление» было установлено и у других птиц, а в постнатальном периоде также у млекопитающих и ряда рыб.
Укажем еще на работы Готтлиба, который показал на утятах, что если эмбрион не подвергается в последние дни инкубации соответствующим акустическим воздействиям, то установление звуковых контактов между вылупившимся птенцом и материнской особью будет значительно затруднено. Изучая затем реакцию следования у нескольких видов уток (крякв, пекинских уток, мандаринок), Готтлиб установил, что наиболее активно утята следовали за макетом (чучелом) при видотипичной визуально-акустической стимуляции, хуже - при одной лишь акустической, т.е. без чучела, и еще хуже при одной оптической, т.е. при неозвученном чучеле. Вместе с тем реакция следования значительно ослабевала, когда видотипичные визуальные или акустические компоненты (позывные звуки и внешний вид чучела) подменялись невидотипичными (т.е. принадлежащими другому виду уток). Канадский ученый Р.М.Эванс в опытах над чайками показал, что искажения в реакциях вылупившихся птенцов (предпочитание криков другого вида) могут обусловливаться отсутствием соответствующего эмбрионального опыта, а именно когда эмбрион лишен возможности слышать крики птиц своего вида.
Итак, как правило, вылупившиеся птенцы реагируют четко избирательно на видотипичные крики, т.е. предпочитают ориентироваться по «натуральным» акустическим раздражителям с большой биологической валентностью. Эти реакции основаны на врожденном узнавании (и предпочтении) видотипичных компонентов коммуникации, которое дополняется и уточняется облигатным выучиванием индивидуальных компонентов общения в процессе пре- и особенно постнатального запечатления. Следовательно, при постнатальном формировании акустического общения между птенцами и их родителями возможность индивидуального (не врожденного!) опознавания особей обеспечивается ранним облигатным научением. Принадлежность же последних к одному и тому же виду устанавливается птенцами на основе врожденного узнавания.
На основе данных, полученных рядом ученых, сейчас можно, очевидно, считать установленным, что и другие функции пения птиц формируются в большой степени благодаря постнатальному облигатному научению. Например, у овсянки песня молодого самца приобретает сексуальную, а затем и территориальную функцию только после того, как он научается петь «полную» песню, т.е. модифицировать ювенильную «подпесню», придать ей специфическую структуру и видоизменить ритм.
Большую роль играет при этом подражание, на что, в частности, еще в середине 50-х годов указывал Торп, отмечая, что птицы учатся непосредственно по звуковым сигналам других птиц.
Выше уже рассматривались явления подражания и говорилось о необходимости выделить как отдельные категории облигатное и факультативное имитационное научение. Теперь должно быть ясно, что облигатное подражание, будучи особым проявлением облигатного научения, характерно прежде всего для раннего постнатального периода онтогенеза. В области звукового общения это можно хорошо проиллюстрировать примерами из многочисленных полевых экспериментов, осуществленных советскими орнитологами К.А. и К.К.Вилкс путем перемещения яиц и птенцов в гнезда других видов (еще раньше подобные эксперименты проводились Промптовым). Изучив поведение приблизительно 4000 пересаженных воробьиных птиц (преимущественно мухоловок-пеструшек), эти исследователи установили, что птенцы в исключительно широком диапазоне перенимают песни «приемных родителей»: более 80% самцов, выросших в гнездах других видов, имитировали песню вида, к которому принадлежала особь, воспитавшая птенца (горихвостки, пеночки-трещотки, большой синицы). Вместе с тем при имитации чужой песни воспроизводились и элементы песни собственного вида. Понятно, что в нормальных условиях акустическое поведение птенца формируется путем облигатного имитационного научения звукам, издаваемым кормящей птицей того же вида.
Примечательно, что перенимание чужих звуков происходит в относительно короткий период начиная с вылета птенца из гнезда. Это доказывается тем, что чужие звуки имитировали и птицы, пересаженные незадолго перед вылетом, но если, наоборот, птенец воспитывался с самого начала в чужом гнезде, но перед вылетом пересаживался обратно к своему виду, чужое воспитание не сказывалось на акустических реакциях такой птицы и ее пение оставалось видотипичным. Таким образом, здесь определенно можно говорить о наличии сенсибильного периода, что опять-таки доказывает, что мы имеем в этом случае дело с облигатным научением, точнее, облигатным имитационным научением.
Раннее формирование общения млекопитающих
У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет - в зависимости от степени зрелорождения, но чаще всего в первые часы жизни новорожденного. Детеныш одногорбого верблюда, например, издает первые звуки еще во время самих родов, а уже час спустя он в состоянии воспроизвести почти все звуки, свойственные его виду, поэтому уже с момента рождения начинается интенсивный акустический контакт между ним и его матерью.
Американским исследователем онтогенеза поведения животных Дж. П. Скоттом и рядом других ученых было доказано, что у собак и других видов псовых существует сенсибильный период, в течение которого только и возможно такое установление контактов с материнской особью и остальными щенками данного выводка. Очевидно, при этом процесс запечатления имеет комплексный характер, поскольку включает в себя и половое запечатление. На это указывает и то обстоятельство, что общение между матерью и щенками, равно как между последними и другими представителями того же вида, может быть по форме идентично половому общению (М. В. Фокс). Так же, по свидетельству ряда ученых, обстоит дело и у обезьян.
Большое значение для формирования общения имеет положительная тигмотаксисная реакция детенышей, выражающаяся в активном установлении физического контакта с поверхностью матери и собратьев, в прижимании к ним. Об этом свидетельствуют, в частности, эксперименты, проведенные на детенышах обезьян Г. Харлоу и его сотрудниками, применявшими разного рода макеты материнских особей (проволочные, тряпочные, обогревамые, холодные и пр.). Было показано, что детеныши предпочитают запечатленный холодный макет даже обогреваемому. Сходная тигмотаксисная реакция, как показали английский этолог В. Слэкин и другие исследователи, присуща также цыплятам и другим птенцам.
Скотт пришел к выводу, что у млекопитающих в раннем постнатальном онтогенезе существуют сенсибильные периоды трех типов: для процессов научения, стимуляции физиологических процессов и формирования общения. Исследования многих ученых показали, что если в течение сенсибильного периода последнего типа детеныш не имеет возможности установить связи с особями своего вида, он впоследствии может оказаться совершенно неспособным к общению с себе подобными и во всяком случае будет испытывать большие затруднения в коммуникативном, а также воспроизводительном поведении. То, что формирование первичных коммуникативных связей совершается путем запечатления, подтверждается и тем, что оно, как доказано рядом экспериментаторов, не зависит от пищевого или иного внешнего подкрепления.
Не менее четко выражено у млекопитающих и «обратное запечатление» - запечатление индивидуальных признаков детенышей у родителей. Так, например, овцы, козы и другие копытные научаются распознавать эти признаки непосредственно после рождения их детенышей и после этого уже не подпускают к соскам чужих. Ведущее значение имеет здесь, вероятно, ольфакторное опознавание, т.е. запоминание индивидуального запаха потомства, хотя и оптические признаки могут играть существенную роль. У детеныша, как показали в эксперименте с чернохвостыми оленями Д. и Х.Мюллер-Шварце, также происходит запечатление индивидуального запаха матери или, в эксперименте, заменяющей модели, причем в отношении последней вполне осуществляются акты общения, проявляемые в норме по отношению к подлинной матери.
Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда, коммуникативные компоненты поведения уже неоднократно встречались нами при ознакомлении с разными аспектами онтогенеза поведения, особенно в процессах запечатления, одним из определяющих факторов которого и является общение.
У птиц, как было уже показано (в частности, в предыдущей главе на кайрах), установление акустических контактов между родительской особью и ее потомством начинается уже на эмбриональной стадии развития птенцов путем пренатального облигатного научения (эмбрионального запечатления). У кайр, например, этот процесс происходит в форме установления акустико-кинестетических связей. В результате этого запечатления птенец подбегает к источнику запечатленного звука. Эта реакция обнаруживается даже у только что вылупившегося птенца: услышав запечатленный звук, он подбегает к родительской особи (в эксперименте к замещающему ее громкоговорителю), издает ответный крик и залезает под нее (или под аппарат). Если же из громкоговорителя после этого раздается другой звук (чужой зов), то птенец немедленно выскакивает из-под него и прячется в укрытии (опыты Чанц). Таким образом, коммуникативный компонент врожденного пускового механизма достраивается в данном случае в ходе эмбрионального запечатления.
Вместе стем и у родительской особи происходит запечатление индивидуальных особенностей голоса еще не вылупившегося птенца, что обеспечивает индивидуальное опознавание последнего уже к моменту вылупления. Подобное «обратное запечатление» было установлено и у других птиц, а в постнатальном периоде также у млекопитающих и ряда рыб.
Укажем еще на работы Готтлиба, который показал на утятах, что если эмбрион не подвергается в последние дни инкубации соответствующим акустическим воздействиям, то установление звуковых контактов между вылупившимся птенцом и материнской особью будет значительно затруднено. Изучая затем реакцию следования у нескольких видов уток (крякв, пекинских уток, мандаринок), Готтлиб установил, что наиболее активно утята следовали за макетом (чучелом) при видотипичной визуально-акустической стимуляции, хуже - при одной лишь акустической, т.е. без чучела, и еще хуже при одной оптической, т.е. при неозвученном чучеле. Вместе с тем реакция следования значительно ослабевала, когда видотипичные визуальные или акустические компоненты (позывные звуки и внешний вид чучела) подменялись невидотипичными (т.е. принадлежащими другому виду уток). Канадский ученый Р.М.Эванс в опытах над чайками показал, что искажения в реакциях вылупившихся птенцов (предпочитание криков другого вида) могут обусловливаться отсутствием соответствующего эмбрионального опыта, а именно когда эмбрион лишен возможности слышать крики птиц своего вида.
Итак, как правило, вылупившиеся птенцы реагируют четко избирательно на видотипичные крики, т.е. предпочитают ориентироваться по «натуральным» акустическим раздражителям с большой биологической валентностью. Эти реакции основаны на врожденном узнавании (и предпочтении) видотипичных компонентов коммуникации, которое дополняется и уточняется облигатным выучиванием индивидуальных компонентов общения в процессе пре- и особенно постнатального запечатления. Следовательно, при постнатальном формировании акустического общения между птенцами и их родителями возможность индивидуального (не врожденного!) опознавания особей обеспечивается ранним облигатным научением. Принадлежность же последних к одному и тому же виду устанавливается птенцами на основе врожденного узнавания.
На основе данных, полученных рядом ученых, сейчас можно, очевидно, считать установленным, что и другие функции пения птиц формируются в большой степени благодаря постнатальному облигатному научению. Например, у овсянки песня молодого самца приобретает сексуальную, а затем и территориальную функцию только после того, как он научается петь «полную» песню, т.е. модифицировать ювенильную «подпесню», придать ей специфическую структуру и видоизменить ритм.
Большую роль играет при этом подражание, на что, в частности, еще в середине 50-х годов указывал Торп, отмечая, что птицы учатся непосредственно по звуковым сигналам других птиц.
Выше уже рассматривались явления подражания и говорилось о необходимости выделить как отдельные категории облигатное и факультативное имитационное научение. Теперь должно быть ясно, что облигатное подражание, будучи особым проявлением облигатного научения, характерно прежде всего для раннего постнатального периода онтогенеза. В области звукового общения это можно хорошо проиллюстрировать примерами из многочисленных полевых экспериментов, осуществленных советскими орнитологами К.А. и К.К.Вилкс путем перемещения яиц и птенцов в гнезда других видов (еще раньше подобные эксперименты проводились Промптовым). Изучив поведение приблизительно 4000 пересаженных воробьиных птиц (преимущественно мухоловок-пеструшек), эти исследователи установили, что птенцы в исключительно широком диапазоне перенимают песни «приемных родителей»: более 80% самцов, выросших в гнездах других видов, имитировали песню вида, к которому принадлежала особь, воспитавшая птенца (горихвостки, пеночки-трещотки, большой синицы). Вместе с тем при имитации чужой песни воспроизводились и элементы песни собственного вида. Понятно, что в нормальных условиях акустическое поведение птенца формируется путем облигатного имитационного научения звукам, издаваемым кормящей птицей того же вида.
Примечательно, что перенимание чужих звуков происходит в относительно короткий период начиная с вылета птенца из гнезда. Это доказывается тем, что чужие звуки имитировали и птицы, пересаженные незадолго перед вылетом, но если, наоборот, птенец воспитывался с самого начала в чужом гнезде, но перед вылетом пересаживался обратно к своему виду, чужое воспитание не сказывалось на акустических реакциях такой птицы и ее пение оставалось видотипичным. Таким образом, здесь определенно можно говорить о наличии сенсибильного периода, что опять-таки доказывает, что мы имеем в этом случае дело с облигатным научением, точнее, облигатным имитационным научением.
Раннее формирование общения млекопитающих
У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет - в зависимости от степени зрелорождения, но чаще всего в первые часы жизни новорожденного. Детеныш одногорбого верблюда, например, издает первые звуки еще во время самих родов, а уже час спустя он в состоянии воспроизвести почти все звуки, свойственные его виду, поэтому уже с момента рождения начинается интенсивный акустический контакт между ним и его матерью.
Американским исследователем онтогенеза поведения животных Дж. П. Скоттом и рядом других ученых было доказано, что у собак и других видов псовых существует сенсибильный период, в течение которого только и возможно такое установление контактов с материнской особью и остальными щенками данного выводка. Очевидно, при этом процесс запечатления имеет комплексный характер, поскольку включает в себя и половое запечатление. На это указывает и то обстоятельство, что общение между матерью и щенками, равно как между последними и другими представителями того же вида, может быть по форме идентично половому общению (М. В. Фокс). Так же, по свидетельству ряда ученых, обстоит дело и у обезьян.
Большое значение для формирования общения имеет положительная тигмотаксисная реакция детенышей, выражающаяся в активном установлении физического контакта с поверхностью матери и собратьев, в прижимании к ним. Об этом свидетельствуют, в частности, эксперименты, проведенные на детенышах обезьян Г. Харлоу и его сотрудниками, применявшими разного рода макеты материнских особей (проволочные, тряпочные, обогревамые, холодные и пр.). Было показано, что детеныши предпочитают запечатленный холодный макет даже обогреваемому. Сходная тигмотаксисная реакция, как показали английский этолог В. Слэкин и другие исследователи, присуща также цыплятам и другим птенцам.
Скотт пришел к выводу, что у млекопитающих в раннем постнатальном онтогенезе существуют сенсибильные периоды трех типов: для процессов научения, стимуляции физиологических процессов и формирования общения. Исследования многих ученых показали, что если в течение сенсибильного периода последнего типа детеныш не имеет возможности установить связи с особями своего вида, он впоследствии может оказаться совершенно неспособным к общению с себе подобными и во всяком случае будет испытывать большие затруднения в коммуникативном, а также воспроизводительном поведении. То, что формирование первичных коммуникативных связей совершается путем запечатления, подтверждается и тем, что оно, как доказано рядом экспериментаторов, не зависит от пищевого или иного внешнего подкрепления.
Не менее четко выражено у млекопитающих и «обратное запечатление» - запечатление индивидуальных признаков детенышей у родителей. Так, например, овцы, козы и другие копытные научаются распознавать эти признаки непосредственно после рождения их детенышей и после этого уже не подпускают к соскам чужих. Ведущее значение имеет здесь, вероятно, ольфакторное опознавание, т.е. запоминание индивидуального запаха потомства, хотя и оптические признаки могут играть существенную роль. У детеныша, как показали в эксперименте с чернохвостыми оленями Д. и Х.Мюллер-Шварце, также происходит запечатление индивидуального запаха матери или, в эксперименте, заменяющей модели, причем в отношении последней вполне осуществляются акты общения, проявляемые в норме по отношению к подлинной матери.