Страница:
Наиболее полноценная, четкая передача информации достигается, однако, тогда, когда появляются специальные двигательные элементы, выделившиеся из «обычных», «утилитарных» форм поведения, утративших в ходе эволюции свою первичную «рабочую», «механическую» функцию и приобретшие чисто сигнальное значение.
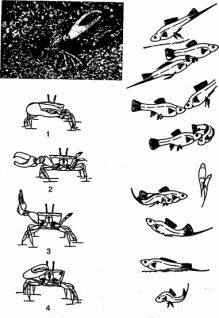

Рис. 10. Сигнальные позы и телодвижения у манящего краба (размахивание самцом клешней при «ухаживании» за самкой), рыбы (меченосца), полевого воробья и лошади (по Крейну, Хеменсу, Блумеи Хассенберг)
Первичные движения получили в этологии название «автохтонных» движений; вторичные, приобретшие новую, в данном случае сигнальную, функцию, - «аллохтонных». В таком случае сам рисунок стереотипно, у всех представителей данного вида одинаково выполняемого движения является условным выражением определенной биологической ситуации (биологически значимого изменения в среде или внутреннего состояния животного). Такие видотипичные стереотипные движения с четкой информативной функцией получили в этологии название «ритуализованных движений». Ритуализация характеризуется условностью выполняемых действий, которые служат лишь для передачи определенного, причем совершенно конкретного информативного содержания. Эти действия генетически жестко фиксированы, выполняются с максимальной стереотипностью, т.е., по существу, одинаково всеми особями данного вида, и поэтому относятся к типичным инстинктивным движениям. По этой причине все животные данного вида и в состоянии «правильно» понимать значение подобных оптических (или акустических) сигналов общения. Это типичный пример максимальной ригидности, консерватизма в поведении животных.
Ритуализованные движения выполняются в строгой последовательности в форме более или менее сложных видотипичных «церемониалов» или «ритуалов», причем, как правило, в виде «диалога» двух животных. Чаще всего они встречаются в сферах размножения (брачные игры) и борьбы («мнимая борьба») и отображают внутреннее состояние особи и ее физические и психические качества. Наряду с врожденными существуют и благоприобретаемые формы общения, о которых пойдет речь позже.
Все формы общения играют существенную роль в жизни высших беспозвоночных и позвоночных, обеспечивая согласованность действий особей. Без коммуникации с помощью различных химических, оптических, акустических, тактильных и других сигналов у этих животных невозможно даже сближение и контактирование самцов и самок, а, следовательно, и продолжение рода. Особое значение приобретает общение особей одного, реже разных видов в условиях их совместной, групповой жизни как основа группового поведения.
Как уже говорилось, групповое поведение появляется (по меньшей мере в развитых формах) вместе с общением только у высших беспозвоночных и выражается в согласованных совместных действиях животных, живущих в сообществах. В отличие от скоплений животных, возникающих в результате идентичной положительной реакции на благоприятные внешние условия, сообщества характеризуются относительно постоянным составом его членов и определенной внутренней структурой, регулирующей отношения между ними. Эта структура нередко принимает форму иерархических систем соподчинения всех членов сообщества друг другу, чем обеспечивается сплоченность последнего и его эффективное функционирование как единого целого. Не меньшее значение имеет в этом отношении выделение вожаков, или «лидеров», регулирование отношений между молодыми и старыми животными, совершенствование ухода за потомством и т.д.
Психический компонент инстинктивного поведения
Глава 3 НАУЧЕНИЕ
Анализ инстинктивного поведения приводит к выводу, что все богатство и разнообразие полноценного психического отражения связаны с научением, накоплением индивидуального опыта. Поэтому эта категория поведения будет постоянно в центре нашего внимания и мы еще не раз обратимся к разным ее аспектам, познакомимся с разными формами научения. Здесь же мы рассмотрим лишь некоторые общие положения.
При изучении процессов научения необходимо исходить из того, что формирование поведения представляет собой процесс конкретного воплощения в жизни особи опыта вида, накопленного и закрепленного в процессе эволюции. Именно этим, как уже отмечалось, определяется переплетение врожденных и индивидуально приобретаемых компонентов поведения. Видовой опыт передается из поколения в поколение в генетически фиксированном виде в форме врожденных, инстинктивных компонентов поведения, поэтому «закодированное» таким образом поведение всегда является видотипичным.
Но это не означает, что справедливо и обратное, т.е. что каждая видотипичная форма поведения по своему происхождению обязательно является врожденной, инстинктивной. Существуют такие формы научения, которые внешне очень напоминают инстинктивное поведение, но тем не менее представляют собой не что иное, как результат накопления индивидуального опыта, правда, в жестких видотипичных рамках. Это прежде всего формы так называемого облигатного научения, которым, по Г. Темброку, обозначается индивидуальный опыт, необходимый для выживания всем представителям данного вида независимо от частных условий жизни особи. Относящиеся сюда формы научения будут рассмотрены при обсуждении онтогенеза поведения.
В противоположность облигатному факультативное научение, согласно Темброку, включает в себя все формы сугубо индивидуального приспособления к особенностям тех конкретных условий, в которых живет данная особь. Понятно, что эти условия не могут быть одинаковыми для всех представителей данного вида. Служа, таким образом, максимальной конкретизации видового поведения в частных условиях среды обитания вида, факультативное научение является наиболее гибким, лабильным компонентом поведения животных. О ряде важных аспектов факультативного научения также будет говориться более подробно в связи с онтогенезом поведения (ч. II, гл. 3). Но следует сразу оговорить, что лабильность факультативного научения неодинакова в разных формах его проявления. Конкретизация видового опыта путем вплетения в инстинктивное поведение элементов индивидуального опыта совершается на всех, даже ригидных, этапах поведенческого акта, хотя, как мы уже знаем, в разной степени. Так, английский этолог Рахайнд указывает на модификацию инстинктивного поведения процессами научения путем изменения сочетания раздражителей, обусловливающих определенный инстинктивный акт, выделения (или усиления) определенных направляющих ключевых раздражителей и т.п.
При этом существенно, что модификации могут иметь место как в сенсорной, так и в эффекторной сфере, хотя чаще всего охватывают обе сферы одновременно. В эффекторной сфере включение элементов научения в инстинктивные действия происходит чаще всего в виде перекомбинации врожденных двигательных элементов, но могут возникнуть и новые двигательные координации. Особенно часто это происходит на более ранних этапах онтогенеза. Примером возникновения таких новых двигательных координации может служить подражательное пение птиц. У высших животных, как мы еще увидим, вновь выученные движения эффекторов, особенно конечностей, играют особенно большую роль в познавательной деятельности, в интеллектуальном поведении.
Что же касается «встраивания» индивидуально-изменчивых компонентов в инстинктивные действия в сенсорной сфере, то это существенно расширяет возможности ориентации животного благодаря овладению новыми сигналами. При этом в отличие от ключевых раздражителей, которые являются для животного изначально биологически значимыми, раздражители, которыми животное руководствуется при научении, первоначально являются для него индифферентными (или, точнее, почти таковыми). Лишь по мере того как животное запоминает их в ходе накопления индивидуального опыта, они приобретают для него сигнальное значение. Таким образом, процесс научения характеризуется избирательным вычленением некоторых «биологически нейтральных» свойств компонентов среды, в результате чего эти свойства приобретают биологическую значимость.
Сейчас мы уже знаем, что этот процесс не сводится просто к образованию условных рефлексов. Дело обстоит значительно сложнее, прежде всего благодаря поистине активному избирательному отношению организма к элементам окружающей среды.
Основой этому служат многообразные динамические процессы в центральной нервной системе, преимущественно в высших ее отделах, благодаря которым осуществляется афферентный синтез раздражений, обусловленные внешними и внутренними факторами. Эти раздражения сопоставляются с ранее воспринятой информацией, хранимой в памяти. В результате формируется готовность к выполнению тех или иных ответных действий, что и проявляется в инициативном, избирательном характере поведения животного по отношению к компонентам среды.
Следующий затем анализ результатов действий животного (по принципу обратной связи) служит основой для нового афферентного синтеза. В результате в дополнение к врожденным, видовым программам поведения в центральной нервной системе постоянно формируются новые, благоприобретаемые, индивидуальные программы, на которых зиждутся процессы научения. Физиологические механизмы этого непрерывного программирования поведения были особенно детально проанализированы П. К. Анохиным и получили свое отражение в его концепции «акцептора результатов действия».
Весьма важным является здесь то, что животное не обречено на пассивное «слепое повиновение» воздействиям среды, а само строит свои отношения с компонентами среды и обладает при этом известной «свободой действия», точнее, свободой поиска и выбора.
Итак, формирование эффективных программ предстоящих действий является результатом комплексных процессов сопоставления и оценки внутренних и внешних раздражений, видового и индивидуального опыта, регистрации параметров совершаемых действий и проверки их результатов. Это и составляет основу научения.
Как мы уже знаем, реализация видового опыта в индивидуальном поведении в наибольшей степени нуждается в процессах научения на начальных этапах поискового поведения. Ведь именно на этих первых этапах каждого поведенческого акта поведение животного является наиболее неопределенным, именно здесь от него требуется максимальная индивидуальная ориентация среди разнородных и переменчивых компонентов среды, и особенно важен быстрый выбор наиболее эффективных способов действия, чтобы быстрее и наилучшим образом достигнуть завершающей фазы поведенческого акта. При этом животное может положиться только на собственный опыт, ибо реакции на единичные, случайные признаки каждой конкретной ситуации, в которой может оказаться любой представитель вида, конечно, не могли быть запрограммированы в процессе эволюции.
Но поскольку без включения благоприобретаемых элементов в инстинктивное поведение последнее неосуществимо, само это включение также наследственно закреплено, как и инстинктивные компоненты поведения. Другими словами, диапазон научения также является строго видотипичным. Представитель любого вида не может научиться «чему угодно», а только тому, что способствует продвижению к завершающим фазам видотипичных поведенческих актов. Существуют, следовательно, видотипичные, генетически фиксированные «лимиты» способности к научению.
Рамки диспозиции к научению не только соответствуют реальным условиям жизни, но у высших животных они подчас и шире, чем того требуют эти условия. Поэтому высшие животные обладают значительными резервами лабильности поведения, потенциальными возможностями индивидуального приспособления к экстремальным ситуациям. У низших животных эти возможности резко сужены, как и вообще способность к научению не достигает у них столь высокого уровня, как у высших животных. Поэтому широта диапазонов способности к научению, наличие указанных резервных возможностей являются показателями высоты психического уровня животных. Чем шире эти диапазоны и чем больше потенциальных возможностей к научению сверх обычной нормы, к накоплению и применению индивидуального опыта в необычных условиях, тем больше и лучше может быть скорректировано инстинктивное поведение, тем более лабильной будет поисковая фаза, тем более пластичным будет все поведение животного и тем выше его общий психический уровень.
Это процесс диалектического взаимодействия, при котором причина и следствие постоянно меняются местами: усложнение в процессе эволюции инстинктивного поведения требует расширения диапазона способности к научению; усиленное же включение элементов научения в инстинктивное поведение делает в результате последнее более пластичным, т.е. приводит к качественному обогащению этого поведения, поднимает его на более высокую ступень. В результате поведение эволюционирует как единое целое.
Эти эволюционные преобразования охватывают как содержание врожденных программ поведения, так и возможности их обогащения индивидуальным опытом, научением. На низших этапах эволюции животного мира эти возможности ограничиваются явлениями привыкания, тренировки и суммационными процессами в сенсорной сфере (см. ч. III). Появляющиеся у низших беспозвоночных временные связи еще весьма неустойчивы, затем они становятся все более прочными, достигая полного развития в разного рода навыках. При привыкании постепенно исчезают реакции на определенное многократно повторяющееся раздражение, не сопровождающееся биологически значимым воздействием на животное. Противоположным явлением можно считать тренировку, при которой инстинктивное действие совершенствуется путем накопления индивидуального опыта.
Разница в поведении между низшими и высшими животными состоит не в том, что более примитивные формы поведения сменяются более сложными, а в том, что к первым добавляются вторые, существенно обогащая и изменяя их, в результате чего элементарные формы поведения приобретают значительную сложность и становятся более вариабельным. Так привыкание, с элементарной формой которого мы еще познакомимся на примере поведения простейших, выступает у млекопитающих в весьма сложных проявлениях. Например, как показали опыты, проведенные Хайндом совместно со Скаурсом, на мышах, реакции этих животных на многократно повторяемые, но ничем не подкрепленные акустические стимулы ослабевают с неодинаковой быстротой. Но эти различия в привыкании обусловливаются не только различиями в интенсивности стимулов (как это имеет место у низших животных), но прежде всего вариабельностью самого процесса привыкания.
Основным качественно новым компонентом научения, дополняющим и обогащающим первичные примитивные формы поведения, является навык, который, однако, на разных эволюционных уровнях тоже проявляется в существенно различающихся формах.
Навык
Рис. 10. Сигнальные позы и телодвижения у манящего краба (размахивание самцом клешней при «ухаживании» за самкой), рыбы (меченосца), полевого воробья и лошади (по Крейну, Хеменсу, Блумеи Хассенберг)
Первичные движения получили в этологии название «автохтонных» движений; вторичные, приобретшие новую, в данном случае сигнальную, функцию, - «аллохтонных». В таком случае сам рисунок стереотипно, у всех представителей данного вида одинаково выполняемого движения является условным выражением определенной биологической ситуации (биологически значимого изменения в среде или внутреннего состояния животного). Такие видотипичные стереотипные движения с четкой информативной функцией получили в этологии название «ритуализованных движений». Ритуализация характеризуется условностью выполняемых действий, которые служат лишь для передачи определенного, причем совершенно конкретного информативного содержания. Эти действия генетически жестко фиксированы, выполняются с максимальной стереотипностью, т.е., по существу, одинаково всеми особями данного вида, и поэтому относятся к типичным инстинктивным движениям. По этой причине все животные данного вида и в состоянии «правильно» понимать значение подобных оптических (или акустических) сигналов общения. Это типичный пример максимальной ригидности, консерватизма в поведении животных.
Ритуализованные движения выполняются в строгой последовательности в форме более или менее сложных видотипичных «церемониалов» или «ритуалов», причем, как правило, в виде «диалога» двух животных. Чаще всего они встречаются в сферах размножения (брачные игры) и борьбы («мнимая борьба») и отображают внутреннее состояние особи и ее физические и психические качества. Наряду с врожденными существуют и благоприобретаемые формы общения, о которых пойдет речь позже.
Все формы общения играют существенную роль в жизни высших беспозвоночных и позвоночных, обеспечивая согласованность действий особей. Без коммуникации с помощью различных химических, оптических, акустических, тактильных и других сигналов у этих животных невозможно даже сближение и контактирование самцов и самок, а, следовательно, и продолжение рода. Особое значение приобретает общение особей одного, реже разных видов в условиях их совместной, групповой жизни как основа группового поведения.
Как уже говорилось, групповое поведение появляется (по меньшей мере в развитых формах) вместе с общением только у высших беспозвоночных и выражается в согласованных совместных действиях животных, живущих в сообществах. В отличие от скоплений животных, возникающих в результате идентичной положительной реакции на благоприятные внешние условия, сообщества характеризуются относительно постоянным составом его членов и определенной внутренней структурой, регулирующей отношения между ними. Эта структура нередко принимает форму иерархических систем соподчинения всех членов сообщества друг другу, чем обеспечивается сплоченность последнего и его эффективное функционирование как единого целого. Не меньшее значение имеет в этом отношении выделение вожаков, или «лидеров», регулирование отношений между молодыми и старыми животными, совершенствование ухода за потомством и т.д.
Психический компонент инстинктивного поведения
Психический компонент инстинктивного поведения, точнее, психическое отражение при инстинктивных действиях животных необходимо изучать на завершающей фазе поведенческого акта и в какой-то степени на непосредственно примыкающем к ней последнем этапе поисковой фазы, поскольку здесь элементы научения играют наименьшую роль. При этом надо сказать, что в свете современных данных и концепций этологии этот вопрос еще специально не изучался и поэтому приходится здесь ограничиваться лишь некоторыми общими соображениями.
Мы видели, что по мере приближения к завершающей фазе движения животного становятся все более ограниченными, однообразными, стереотипными, и есть все основания полагать, что в такой же мере отражение окружающего мира становится все более бедным, ограниченным. Если на первых этапах поискового поведения животное в поиске ключевых раздражителей руководствуется многими разнообразными ориентирами в окружающей среде, то по мере сужения действия внешних агентов, направляющих поведение животного, ориентация все больше осуществляется только по направляющим ключевым раздражителям, а в завершающей фазе производится уже исключительно по ним.
Вместе с тем мы теперь уже знаем, что как направляющие, так тем более и пусковые ключевые раздражители представляют собой элементарные физические и химические признаки, воспринимаемые животным «в отрыве» от объекта, которому они присущи. Следовательно, животное получает на завершающей фазе лишь весьма неполную, «однобокую» информацию о некоторых - причем несущественных - внешних признаках объектов инстинктивных действий. Правда, эти признаки являются постоянными и в этом отношении характерными для этих объектов.
Однако, что для нас сейчас важно, животное, реагируя на эти признаки, в сущности, не получает сведений о самом объекте (неодушевленном предмете или другом живом существе): признаки - ключевые раздражители - являются только ориентирами, направляющими действия животного на этот объект, носитель этих ориентиров.
Это с совершенной очевидностью вытекает из приведенных выше экспериментов, в которых подлинные объекты инстинктивных действий заменялись весьма абстрактными макетами. Вспомним, что в этих условиях удалось получить даже сверхоптимальные «преувеличенные» инстинктивные реакции. Самец колюшки в завершающей фазе борьбы за гнездовой участок не видит разницы между живой рыбой-соперником и куском жести или пластмассы. Он видит только «красное», и этого ему в данной ситуации вполне достаточно, чтобы среагировать на это «красное», которое в данном случае, кстати, выступает одновременно как направляющий и пусковой ключевой раздражитель.
Следовательно, здесь налицо первично генерализованная реакция на общий элементарный признак (точнее, на совокупность нескольких таких признаков, поскольку в природных условиях действуют не одно только «красное», но и некоторые другие ключевые раздражители). Этот признак воспринимается недифференцированно, как простое ощущение (точнее, воспринимаются суммы ощущений). Об этом свидетельствует отсутствие реакций на форму объекта - носителя признака, да и вообще на объект (предмет или рыбу) как таковой. Ясно, что здесь обнаруживается очень бедное, крайне поверхностное и ограниченное отражение окружающего мира, низшая форма психического отражения. Неинтересно, что такое отражение, свойственное низшим представителям животного мира, в полной мере сохранилось и у высших животных - позвоночных и продолжает у них играть главную роль в поддержании жизнедеятельности особи и продолжении рода.
О том, как это понимать, уже говорилось выше. Эволюция животной жизни могла идти только по пути отбора как можно более простых, но и наиболее постоянных ориентиров (или, что не меняет существа дела, создания таких специфических ориентиров, когда речь идет о животных одного вида, являющихся носителями ключевых признаков), руководствуясь которыми организмы могли бы успешно, не отвлекаясь другими, случайными, изменчивыми признаками, осуществлять наиважнейшие жизненные функции.
Правда, как явствует, например, из описанных опытов с птенцами дроздов, у высших животных ключевые стимулы представлены и более сложными по своей структуре типами. В указанном примере имеет место реакция на конфигурацию и на пространственные соотношения деталей объектов инстинктивных движений, что уже соответствует элементарным восприятиям. Это, очевидно, одно из важных проявлений прогрессивного развития инстинктивного поведения в ходе эволюции. Однако, как показывают те же эксперименты, и здесь нет подлинного предметного восприятия, ибо иначе птенцы не могли бы спутать свою мать с картонным диском.
Примитивность психического отражения на завершающей фазе инстинктивных действий является следствием бедности самой моторики в этой фазе. Как мы знаем, двигательная активность, направленная на окружающую среду, является источником познания этой среды. Однако столь стереотипные движения со столь ограниченной, специальной функцией, какими являются инстинктивные движения, врожденные двигательные координации, не могут служить сколько-нибудь пригодной основой для познания окружающего мира.
Врожденные двигательные координации (во всяком случае в их типичной форме) и не выполняют эту функцию, а ключевые раздражители, воспринимаемые на завершающей фазе (да и на отдельных этапах поискового поведения), не познаются, а узнаются врожденным образом благодаря наличию врожденных пусковых механизмов, чтобы затем использоваться как ориентиры или - и того меньше - как «кнопки» пуска инстинктивных реакций. Здесь больше ничего нет, кроме, вероятно, положительной или отрицательной эмоциональной окраски ощущений от воспринимаемых стимулов и собственных движений.
Познавательные же функции и вообще все богатство психического отражения приурочены к начальным этапам поискового поведения, где в полной мере действуют процессы научения.
Мы видели, что по мере приближения к завершающей фазе движения животного становятся все более ограниченными, однообразными, стереотипными, и есть все основания полагать, что в такой же мере отражение окружающего мира становится все более бедным, ограниченным. Если на первых этапах поискового поведения животное в поиске ключевых раздражителей руководствуется многими разнообразными ориентирами в окружающей среде, то по мере сужения действия внешних агентов, направляющих поведение животного, ориентация все больше осуществляется только по направляющим ключевым раздражителям, а в завершающей фазе производится уже исключительно по ним.
Вместе с тем мы теперь уже знаем, что как направляющие, так тем более и пусковые ключевые раздражители представляют собой элементарные физические и химические признаки, воспринимаемые животным «в отрыве» от объекта, которому они присущи. Следовательно, животное получает на завершающей фазе лишь весьма неполную, «однобокую» информацию о некоторых - причем несущественных - внешних признаках объектов инстинктивных действий. Правда, эти признаки являются постоянными и в этом отношении характерными для этих объектов.
Однако, что для нас сейчас важно, животное, реагируя на эти признаки, в сущности, не получает сведений о самом объекте (неодушевленном предмете или другом живом существе): признаки - ключевые раздражители - являются только ориентирами, направляющими действия животного на этот объект, носитель этих ориентиров.
Это с совершенной очевидностью вытекает из приведенных выше экспериментов, в которых подлинные объекты инстинктивных действий заменялись весьма абстрактными макетами. Вспомним, что в этих условиях удалось получить даже сверхоптимальные «преувеличенные» инстинктивные реакции. Самец колюшки в завершающей фазе борьбы за гнездовой участок не видит разницы между живой рыбой-соперником и куском жести или пластмассы. Он видит только «красное», и этого ему в данной ситуации вполне достаточно, чтобы среагировать на это «красное», которое в данном случае, кстати, выступает одновременно как направляющий и пусковой ключевой раздражитель.
Следовательно, здесь налицо первично генерализованная реакция на общий элементарный признак (точнее, на совокупность нескольких таких признаков, поскольку в природных условиях действуют не одно только «красное», но и некоторые другие ключевые раздражители). Этот признак воспринимается недифференцированно, как простое ощущение (точнее, воспринимаются суммы ощущений). Об этом свидетельствует отсутствие реакций на форму объекта - носителя признака, да и вообще на объект (предмет или рыбу) как таковой. Ясно, что здесь обнаруживается очень бедное, крайне поверхностное и ограниченное отражение окружающего мира, низшая форма психического отражения. Неинтересно, что такое отражение, свойственное низшим представителям животного мира, в полной мере сохранилось и у высших животных - позвоночных и продолжает у них играть главную роль в поддержании жизнедеятельности особи и продолжении рода.
О том, как это понимать, уже говорилось выше. Эволюция животной жизни могла идти только по пути отбора как можно более простых, но и наиболее постоянных ориентиров (или, что не меняет существа дела, создания таких специфических ориентиров, когда речь идет о животных одного вида, являющихся носителями ключевых признаков), руководствуясь которыми организмы могли бы успешно, не отвлекаясь другими, случайными, изменчивыми признаками, осуществлять наиважнейшие жизненные функции.
Правда, как явствует, например, из описанных опытов с птенцами дроздов, у высших животных ключевые стимулы представлены и более сложными по своей структуре типами. В указанном примере имеет место реакция на конфигурацию и на пространственные соотношения деталей объектов инстинктивных движений, что уже соответствует элементарным восприятиям. Это, очевидно, одно из важных проявлений прогрессивного развития инстинктивного поведения в ходе эволюции. Однако, как показывают те же эксперименты, и здесь нет подлинного предметного восприятия, ибо иначе птенцы не могли бы спутать свою мать с картонным диском.
Примитивность психического отражения на завершающей фазе инстинктивных действий является следствием бедности самой моторики в этой фазе. Как мы знаем, двигательная активность, направленная на окружающую среду, является источником познания этой среды. Однако столь стереотипные движения со столь ограниченной, специальной функцией, какими являются инстинктивные движения, врожденные двигательные координации, не могут служить сколько-нибудь пригодной основой для познания окружающего мира.
Врожденные двигательные координации (во всяком случае в их типичной форме) и не выполняют эту функцию, а ключевые раздражители, воспринимаемые на завершающей фазе (да и на отдельных этапах поискового поведения), не познаются, а узнаются врожденным образом благодаря наличию врожденных пусковых механизмов, чтобы затем использоваться как ориентиры или - и того меньше - как «кнопки» пуска инстинктивных реакций. Здесь больше ничего нет, кроме, вероятно, положительной или отрицательной эмоциональной окраски ощущений от воспринимаемых стимулов и собственных движений.
Познавательные же функции и вообще все богатство психического отражения приурочены к начальным этапам поискового поведения, где в полной мере действуют процессы научения.
Глава 3 НАУЧЕНИЕ
Общая характеристика процесса научения
Анализ инстинктивного поведения приводит к выводу, что все богатство и разнообразие полноценного психического отражения связаны с научением, накоплением индивидуального опыта. Поэтому эта категория поведения будет постоянно в центре нашего внимания и мы еще не раз обратимся к разным ее аспектам, познакомимся с разными формами научения. Здесь же мы рассмотрим лишь некоторые общие положения.
При изучении процессов научения необходимо исходить из того, что формирование поведения представляет собой процесс конкретного воплощения в жизни особи опыта вида, накопленного и закрепленного в процессе эволюции. Именно этим, как уже отмечалось, определяется переплетение врожденных и индивидуально приобретаемых компонентов поведения. Видовой опыт передается из поколения в поколение в генетически фиксированном виде в форме врожденных, инстинктивных компонентов поведения, поэтому «закодированное» таким образом поведение всегда является видотипичным.
Но это не означает, что справедливо и обратное, т.е. что каждая видотипичная форма поведения по своему происхождению обязательно является врожденной, инстинктивной. Существуют такие формы научения, которые внешне очень напоминают инстинктивное поведение, но тем не менее представляют собой не что иное, как результат накопления индивидуального опыта, правда, в жестких видотипичных рамках. Это прежде всего формы так называемого облигатного научения, которым, по Г. Темброку, обозначается индивидуальный опыт, необходимый для выживания всем представителям данного вида независимо от частных условий жизни особи. Относящиеся сюда формы научения будут рассмотрены при обсуждении онтогенеза поведения.
В противоположность облигатному факультативное научение, согласно Темброку, включает в себя все формы сугубо индивидуального приспособления к особенностям тех конкретных условий, в которых живет данная особь. Понятно, что эти условия не могут быть одинаковыми для всех представителей данного вида. Служа, таким образом, максимальной конкретизации видового поведения в частных условиях среды обитания вида, факультативное научение является наиболее гибким, лабильным компонентом поведения животных. О ряде важных аспектов факультативного научения также будет говориться более подробно в связи с онтогенезом поведения (ч. II, гл. 3). Но следует сразу оговорить, что лабильность факультативного научения неодинакова в разных формах его проявления. Конкретизация видового опыта путем вплетения в инстинктивное поведение элементов индивидуального опыта совершается на всех, даже ригидных, этапах поведенческого акта, хотя, как мы уже знаем, в разной степени. Так, английский этолог Рахайнд указывает на модификацию инстинктивного поведения процессами научения путем изменения сочетания раздражителей, обусловливающих определенный инстинктивный акт, выделения (или усиления) определенных направляющих ключевых раздражителей и т.п.
При этом существенно, что модификации могут иметь место как в сенсорной, так и в эффекторной сфере, хотя чаще всего охватывают обе сферы одновременно. В эффекторной сфере включение элементов научения в инстинктивные действия происходит чаще всего в виде перекомбинации врожденных двигательных элементов, но могут возникнуть и новые двигательные координации. Особенно часто это происходит на более ранних этапах онтогенеза. Примером возникновения таких новых двигательных координации может служить подражательное пение птиц. У высших животных, как мы еще увидим, вновь выученные движения эффекторов, особенно конечностей, играют особенно большую роль в познавательной деятельности, в интеллектуальном поведении.
Что же касается «встраивания» индивидуально-изменчивых компонентов в инстинктивные действия в сенсорной сфере, то это существенно расширяет возможности ориентации животного благодаря овладению новыми сигналами. При этом в отличие от ключевых раздражителей, которые являются для животного изначально биологически значимыми, раздражители, которыми животное руководствуется при научении, первоначально являются для него индифферентными (или, точнее, почти таковыми). Лишь по мере того как животное запоминает их в ходе накопления индивидуального опыта, они приобретают для него сигнальное значение. Таким образом, процесс научения характеризуется избирательным вычленением некоторых «биологически нейтральных» свойств компонентов среды, в результате чего эти свойства приобретают биологическую значимость.
Сейчас мы уже знаем, что этот процесс не сводится просто к образованию условных рефлексов. Дело обстоит значительно сложнее, прежде всего благодаря поистине активному избирательному отношению организма к элементам окружающей среды.
Основой этому служат многообразные динамические процессы в центральной нервной системе, преимущественно в высших ее отделах, благодаря которым осуществляется афферентный синтез раздражений, обусловленные внешними и внутренними факторами. Эти раздражения сопоставляются с ранее воспринятой информацией, хранимой в памяти. В результате формируется готовность к выполнению тех или иных ответных действий, что и проявляется в инициативном, избирательном характере поведения животного по отношению к компонентам среды.
Следующий затем анализ результатов действий животного (по принципу обратной связи) служит основой для нового афферентного синтеза. В результате в дополнение к врожденным, видовым программам поведения в центральной нервной системе постоянно формируются новые, благоприобретаемые, индивидуальные программы, на которых зиждутся процессы научения. Физиологические механизмы этого непрерывного программирования поведения были особенно детально проанализированы П. К. Анохиным и получили свое отражение в его концепции «акцептора результатов действия».
Весьма важным является здесь то, что животное не обречено на пассивное «слепое повиновение» воздействиям среды, а само строит свои отношения с компонентами среды и обладает при этом известной «свободой действия», точнее, свободой поиска и выбора.
Итак, формирование эффективных программ предстоящих действий является результатом комплексных процессов сопоставления и оценки внутренних и внешних раздражений, видового и индивидуального опыта, регистрации параметров совершаемых действий и проверки их результатов. Это и составляет основу научения.
Как мы уже знаем, реализация видового опыта в индивидуальном поведении в наибольшей степени нуждается в процессах научения на начальных этапах поискового поведения. Ведь именно на этих первых этапах каждого поведенческого акта поведение животного является наиболее неопределенным, именно здесь от него требуется максимальная индивидуальная ориентация среди разнородных и переменчивых компонентов среды, и особенно важен быстрый выбор наиболее эффективных способов действия, чтобы быстрее и наилучшим образом достигнуть завершающей фазы поведенческого акта. При этом животное может положиться только на собственный опыт, ибо реакции на единичные, случайные признаки каждой конкретной ситуации, в которой может оказаться любой представитель вида, конечно, не могли быть запрограммированы в процессе эволюции.
Но поскольку без включения благоприобретаемых элементов в инстинктивное поведение последнее неосуществимо, само это включение также наследственно закреплено, как и инстинктивные компоненты поведения. Другими словами, диапазон научения также является строго видотипичным. Представитель любого вида не может научиться «чему угодно», а только тому, что способствует продвижению к завершающим фазам видотипичных поведенческих актов. Существуют, следовательно, видотипичные, генетически фиксированные «лимиты» способности к научению.
Рамки диспозиции к научению не только соответствуют реальным условиям жизни, но у высших животных они подчас и шире, чем того требуют эти условия. Поэтому высшие животные обладают значительными резервами лабильности поведения, потенциальными возможностями индивидуального приспособления к экстремальным ситуациям. У низших животных эти возможности резко сужены, как и вообще способность к научению не достигает у них столь высокого уровня, как у высших животных. Поэтому широта диапазонов способности к научению, наличие указанных резервных возможностей являются показателями высоты психического уровня животных. Чем шире эти диапазоны и чем больше потенциальных возможностей к научению сверх обычной нормы, к накоплению и применению индивидуального опыта в необычных условиях, тем больше и лучше может быть скорректировано инстинктивное поведение, тем более лабильной будет поисковая фаза, тем более пластичным будет все поведение животного и тем выше его общий психический уровень.
Это процесс диалектического взаимодействия, при котором причина и следствие постоянно меняются местами: усложнение в процессе эволюции инстинктивного поведения требует расширения диапазона способности к научению; усиленное же включение элементов научения в инстинктивное поведение делает в результате последнее более пластичным, т.е. приводит к качественному обогащению этого поведения, поднимает его на более высокую ступень. В результате поведение эволюционирует как единое целое.
Эти эволюционные преобразования охватывают как содержание врожденных программ поведения, так и возможности их обогащения индивидуальным опытом, научением. На низших этапах эволюции животного мира эти возможности ограничиваются явлениями привыкания, тренировки и суммационными процессами в сенсорной сфере (см. ч. III). Появляющиеся у низших беспозвоночных временные связи еще весьма неустойчивы, затем они становятся все более прочными, достигая полного развития в разного рода навыках. При привыкании постепенно исчезают реакции на определенное многократно повторяющееся раздражение, не сопровождающееся биологически значимым воздействием на животное. Противоположным явлением можно считать тренировку, при которой инстинктивное действие совершенствуется путем накопления индивидуального опыта.
Разница в поведении между низшими и высшими животными состоит не в том, что более примитивные формы поведения сменяются более сложными, а в том, что к первым добавляются вторые, существенно обогащая и изменяя их, в результате чего элементарные формы поведения приобретают значительную сложность и становятся более вариабельным. Так привыкание, с элементарной формой которого мы еще познакомимся на примере поведения простейших, выступает у млекопитающих в весьма сложных проявлениях. Например, как показали опыты, проведенные Хайндом совместно со Скаурсом, на мышах, реакции этих животных на многократно повторяемые, но ничем не подкрепленные акустические стимулы ослабевают с неодинаковой быстротой. Но эти различия в привыкании обусловливаются не только различиями в интенсивности стимулов (как это имеет место у низших животных), но прежде всего вариабельностью самого процесса привыкания.
Основным качественно новым компонентом научения, дополняющим и обогащающим первичные примитивные формы поведения, является навык, который, однако, на разных эволюционных уровнях тоже проявляется в существенно различающихся формах.
Навык
Основные особенности навыков
Навык - центральная, важнейшая форма факультативного научения. Иногда термин «навык» употребляется для обозначения научения вообще. Однако, как подчеркивает А. Н. Леонтьев, нельзя называть навыками любые связи, возникающие в индивидуальном опыте, ибо при таком расширенном понимании понятие навыка становится весьма расплывчатым, а тем самым непригодным для строго научного анализа. Уже указывалось на то, что способность к выработке навыков свойственна не всем животным, а проявляется лишь на определенном уровне филогенеза. В результате формирования навыка применяется врожденная двигательная координация (обычного поведения) в новой сигнальной ситуации или возникает новая (приобретенная) двигательная координация. В последнем случае появляются новые, генетически не фиксированные движения, т.е. животное научается что-то делать по-новому. Но в любом случае решающее значение имеет успешность выполняемых движений и их подкрепление положительным результатом. При этом, по существу, неважно, строится ли научение на информации, полученной путем собственного активного поиска раздражителей, либо в ходе общения с другими животными, коммуникаций, куда относятся также случаи подражания или обучения (человеком - животного, взрослой особью - детеныша и т.п.).
Другой важнейший признак навыка, также связанный с необходимостью подкрепления, заключается в том, что он формируется в результате упражнения и нуждается для своего дальнейшего сохранения в тренировке. При тренировке навыки совершенствуются, при отсутствии же ее угасают, разрушаются.
Навыки интенсивно изучаются у животных с помощью разнообразных специальных методов: «лабиринта», «проблемного ящика» (или клетки), «обходного пути» и др. При всех этих методах животное ставится в условия выбора сигналов или способов действия при решении определенной задачи.
Уже описанный выше метод «лабиринта» был введен в практику экспериментального исследования в 1901 г. американским психологом В. С. Смоллом. В настоящее время применяются лабиринты самой различной конструкции (см. рис. 1), но задача, которую приходится решать подопытному животному, всегда одна и та же - как можно быстрее и без захода в тупики достигнуть места подкрепления (чаще всего пищевого). Большинство опытов проводилось с крысами, которые легко справляются с такими задачами. Практически не существует лабиринта, в котором крыса не сумела бы сориентироваться. Особенно интенсивно пользовались лабиринтами, как и проблемными ящиками и клетками (также уже описанными), бихевиористы. Проблемный ящик был усовершенствован и снабжен автоматическими устройствами американским психологом Б. Ф. Скиннером. В качестве подопытных животных при этом также использовались крысы.
В лабиринте животное решает задачу как бы «вслепую», так как реакция (пробежка) вырабатывается без непосредственного контакта с обусловливающим ее стимулом (пищей, гнездом), и более того, животное в начале опыта еще ничего не знает о наличии целевого объекта, а лишь случайно обнаруживает его впоследствии в результате ориентировочно-исследовательских действий. Запоминание этого объекта и пути к нему и служат основой формирования навыка. Если впоследствии животное многократно пробежит это расстояние одним и тем же выученным кратчайшим путем, то этот навык становится стереотипным, автоматизированным. Пластичность поведения в этом случае невелика, а при сильно выраженной стереотипности выученных движений последние подчас приближаются к инстинктивным двигательным стереотипам. Стереотипность вообще характерна для примитивных навыков, при которых формируются ригидные, автоматизированные двигательные реакции. Значительная пластичность проявляется при таких навыках лишь па первых этапах образования этих навыков. (С навыками высшего порядка, отличающимися большой пластичностью, мы познакомимся позже.)
Навык - центральная, важнейшая форма факультативного научения. Иногда термин «навык» употребляется для обозначения научения вообще. Однако, как подчеркивает А. Н. Леонтьев, нельзя называть навыками любые связи, возникающие в индивидуальном опыте, ибо при таком расширенном понимании понятие навыка становится весьма расплывчатым, а тем самым непригодным для строго научного анализа. Уже указывалось на то, что способность к выработке навыков свойственна не всем животным, а проявляется лишь на определенном уровне филогенеза. В результате формирования навыка применяется врожденная двигательная координация (обычного поведения) в новой сигнальной ситуации или возникает новая (приобретенная) двигательная координация. В последнем случае появляются новые, генетически не фиксированные движения, т.е. животное научается что-то делать по-новому. Но в любом случае решающее значение имеет успешность выполняемых движений и их подкрепление положительным результатом. При этом, по существу, неважно, строится ли научение на информации, полученной путем собственного активного поиска раздражителей, либо в ходе общения с другими животными, коммуникаций, куда относятся также случаи подражания или обучения (человеком - животного, взрослой особью - детеныша и т.п.).
Другой важнейший признак навыка, также связанный с необходимостью подкрепления, заключается в том, что он формируется в результате упражнения и нуждается для своего дальнейшего сохранения в тренировке. При тренировке навыки совершенствуются, при отсутствии же ее угасают, разрушаются.
Навыки интенсивно изучаются у животных с помощью разнообразных специальных методов: «лабиринта», «проблемного ящика» (или клетки), «обходного пути» и др. При всех этих методах животное ставится в условия выбора сигналов или способов действия при решении определенной задачи.
Уже описанный выше метод «лабиринта» был введен в практику экспериментального исследования в 1901 г. американским психологом В. С. Смоллом. В настоящее время применяются лабиринты самой различной конструкции (см. рис. 1), но задача, которую приходится решать подопытному животному, всегда одна и та же - как можно быстрее и без захода в тупики достигнуть места подкрепления (чаще всего пищевого). Большинство опытов проводилось с крысами, которые легко справляются с такими задачами. Практически не существует лабиринта, в котором крыса не сумела бы сориентироваться. Особенно интенсивно пользовались лабиринтами, как и проблемными ящиками и клетками (также уже описанными), бихевиористы. Проблемный ящик был усовершенствован и снабжен автоматическими устройствами американским психологом Б. Ф. Скиннером. В качестве подопытных животных при этом также использовались крысы.
В лабиринте животное решает задачу как бы «вслепую», так как реакция (пробежка) вырабатывается без непосредственного контакта с обусловливающим ее стимулом (пищей, гнездом), и более того, животное в начале опыта еще ничего не знает о наличии целевого объекта, а лишь случайно обнаруживает его впоследствии в результате ориентировочно-исследовательских действий. Запоминание этого объекта и пути к нему и служат основой формирования навыка. Если впоследствии животное многократно пробежит это расстояние одним и тем же выученным кратчайшим путем, то этот навык становится стереотипным, автоматизированным. Пластичность поведения в этом случае невелика, а при сильно выраженной стереотипности выученных движений последние подчас приближаются к инстинктивным двигательным стереотипам. Стереотипность вообще характерна для примитивных навыков, при которых формируются ригидные, автоматизированные двигательные реакции. Значительная пластичность проявляется при таких навыках лишь па первых этапах образования этих навыков. (С навыками высшего порядка, отличающимися большой пластичностью, мы познакомимся позже.)