Страница:
Виноградные улитки же выполняют сложные «брачные игры», длящиеся иногда по нескольку часов, во время которых партнеры принимают по отношению друг к другу различные позы, колют друг друга известковыми спикулами («любовными стрелами») и т.д. Только после такой взаимной стимуляции начинается собственно спаривание (перенос сперматофора). У некоторых полихет, например у Platynereis dumerilii, в последние годы также были описаны брачные «танцы», однако, очевидно, еще рано делать выводы о их конкретном значении и о том, имеют ли они какое-то отношение к ритуализованному поведению. Но во всех этих случаях, несомненно, обнаруживаются некие зачатки, предваряющие значительно более сложные формы поведения высших позвоночных.
Пластичность поведения
Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих животных еще небольшую роль. Ассоциативные связи формируются у них с трудом и лишь в ограниченных пределах. Результаты научения сохраняются недолго. У всех кольчатых червей встречается наиболее простая форма научения - привыкание, с которым мы уже встречались у простейших, у которых она является главной, если не единственной формой модификации врожденного, видотипичного поведения. У низших многоклеточных животных врожденные реакции на определенные раздражения также прекращаются после многократного повторения, если не последует адекватного подкрепления этих реакций. Так, например, дождевые черви перестают реагировать на повторное затенение, если оно остается без последствий.
Отмечается привыкание и в сфере пищевого поведения: если многощетинкового червя повторно «кормить» комочками бумаги, смоченными соком его обычной жертвы, он перестает их принимать. Если попеременно с такими комочками давать ему подлинные кусочки пищи, то он в конце концов научается различать их и будет отвергать лишь несъедобную бумагу. Подобные опыты ставились и на кишечнополостных (на полипах, находящихся, как мы уже знаем, на более низком уровне элементарной сенсорной психики). При этом полипы вели себя так же, как морские черви: уже после нескольких (до 5) опытов они отбрасывали несъедобные объекты еще до того, как подносили их к ротовому отверстию. Интересно, что нетренированные щупальца этого не делали, даже находясь рядом с тренированными.
Эти эксперименты интересны тем, что показывают способность низших беспозвоночных отличить по побочным физическим качествам съедобное от несъедобного (по вкусу предлагавшиеся объекты были одинаковыми), что подтверждает наличие истинного психического отражения уже на этом, низшем, уровне филогенетического развития. Ведь здесь производится опосредствованное действие, отличающееся тем, что свойство (или сочетание нескольких свойств), которым животное руководствуется при оценке пригодности объекта к пищевому употреблению, выступает как подлинный сигнал, а чувствительность червя или полипа играет роль посредника между организмом и компонентом среды, от которого непосредственно зависит существование животного.
Более сложное научение путем «проб и ошибок» и формирования новой индивидуальной двигательной реакции можно в элементарной форме обнаружить, начиная уже с плоских червей. Так, планарии, встречая на своем пути полоску наждачной бумаги, сперва останавливаются, но затем все же переползают через нее. Если же сочетать соприкосновение червя с наждачной бумагой с его сотрясением, то планарии перестают переползать через нее и тогда, когда не производится сотрясения. Правда, здесь, очевидно, еще нет подлинной ассоциации между двумя раздражителями (шероховатость и сотрясение); скорее всего, происходит суммация первоначально не очень сильного отрицательного раздражения (шероховатость) с дополнительным отрицательным раздражением (сотрясение) и что, вероятно, влечет за собой общее повышение возбудимости животного.
Все же можно выработать уже у планарии и более сложные реакции, когда один из раздражителей является биологически «нейтральным». В таких случаях можно говорить, очевидно, уже об элементарных процессах подлинного ассоциативного научения. Так, например, Л.Г.Воронин и Н.А.Тушмалова сумели выработать у планарии и кольчатых червей оборонительные и пищевые условные рефлексы. При этом были получены и данные, свидетельствующие об усложнении форм временных связей. Если по поводу плоских червей можно говорить лишь о примитивных неустойчивых условных рефлексах, то у многощетинковых червей обнаруживаются уже стабильные самовосстанавливающиеся после угасания условные рефлексы. Эти различия соответствуют глубоким морфологическим различиям в строении нервной системы этих групп животных и свидетельствуют о существенном прогрессе психической активности у полихет.
Пластичность поведения дождевых червей очень убедительно была показана еще в 1912 г. известным американским зоопсихологом Р.Иерксом. В проведенных им опытах черви должны были в Т-образном лабиринте выбирать определенную сторону, где находилось «гнездо» (в противоположной стороне червь получал электрический удар). Чтобы этому научить червей, потребовалось 120- 180 опытов. (Улитки осваивают такую задачу после 60 опытов и запоминают верное решение в течение 30 дней.) Впоследствии аналогичные опыты с дождевыми червями ставились Л.Геком, И.С.Робинсоном, Р.И.Войтусяком (рис. 34) и другими исследователями. Удавалось также переучивать червей, поменяв «гнездо» и электроды местами. Характерно, что результат научения не меняется и после удаления передних сегментов. Более того, даже лишенный с самого начала этих сегментов, червь оказывается способным научиться правильно ориентироваться в лабиринте.
Подобные факты побудили в свое время В.А. Вагнера говорить о «сегментарной психологии» кольчатых червей и других «членистых», имея при этом в виду, что ганглии каждого сегмента в большой степени обеспечивают автономное выполнение элементарных психических функций. Обезглавленные кольчецы, писал Вагнер, «не теряют способности к спонтанным движениям, за исключением тех лишь, которые стоят в прямой зависимости и связи с органами чувств головы… Обезглавленные черви удерживают свои инстинктивные действия, даже те, в которых голова принимает прямое участие».*
* Вагнер В.А.Сегментарная психология. Новые идеи в биологии. Сборник № 6. Биопсихология. Спб., 1914. С. 123.

Рис. 34. Научение у дождевого червя (опыты Войтусяка)
Все это свидетельствует о том, что на уровне даже высших червей цефализация еще не достигла такого развития, чтобы оказать решающее влияние на все поведенческие акты животного. Все же общая направляющая роль головного мозга достаточно велика уже и на этом уровне. Это проявляется, например, в том, что если разрезать пополам дождевого червя, передняя половина будет закапываться в почву, производя вполне координированные движения, задняя же будет лишь беспорядочно извиваться.
Еще лучше, чем у малощетинковых червей, как уже отмечалось, вырабатываются ассоциативные связи у полихет. В опытах удавалось, например, переделать отрицательную реакцию на освещение в положительную путем сочетания освещения с подкармливанием. В итоге черви стали при освещении выползать из своих домиков и без пищевого подкрепления. Время реакции (между световым раздражением и ответным движением) уменьшилось при этом с 28,9 до 3,5 секунды (опыты М.Купленда).
Обстоятельно изучал способность к научению полихет С.М.Эванс. Применяя наказания электрическим током, этот исследователь довольно легко выработал у подопытных животных отрицательную двигательную реакцию (отказ от вползания в узкий темный коридор). В других опытах червям приходилось проползать Т-образный лабиринт. При правильном повороте они могли в течение 5 минут пребывать в затемненной камере. Особенно хорошо обучались полихеты Nereis virens, которым понадобилось менее 100 сочетаний для почти безошибочного решения задачи и ее прочного запоминания; спустя 48 часов они безошибочно воспроизводили правильный путь через лабиринт (рис. 35).
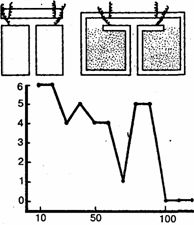
Рис. 35. Научение у Nereis virens (опытЭванса). Вверху даны схемы Т-образных лабиринтов, в которых производились опыты. Внизу - кривая научения (по оси ординат-число ошибок, по оси абсцисс - число опытов)
Общая характеристика высшего уровня элементарной сенсорной психики
Как уже отмечалось, наиболее низкоорганизованные формы многоклеточных беспозвоночных стоят на том же уровне психического развития, что и высшие представители простейших. Но то, что здесь не обнаруживаются существенные различия в поведении, несмотря на глубокие различия в строении, не должно нас удивлять, ибо, как уже говорилось, простейшие олицетворяют собой совершенно особую, рано отклонившуюся филогенетическую ветвь, которая до известных пределов развивалась параллельно ветви низших многоклеточных животных.
Что же касается поведения кольчатых червей, то оно вполне отвечает стадии элементарной сенсорной психики, ибо слагается из движений, ориентированных лишь по отдельным свойствам предметов и явлений (или их сочетаниям), причем это те свойства, которые, по Леонтьеву, оповещают о появлении жизненно важных условий среды, от которых зависит осуществление основных биологических функций животных. Эта ориентация осуществляется, таким образом, на основе одних лишь ощущений. Перцепция, способность к предметному восприятию, еще отсутствует. Не исключено, правда, что у некоторых наземных улиток, как и у упомянутых выше свободно плавающих хищных моллюсков и полихет, уже намечаются зачатки этой способности. Так, виноградная улитка обходит преграду еще до прикосновения к ней, ползет вдоль нее, но только если преграда не слишком велика; если же изображение преграды занимает всю сетчатку, улитка наталкивается на нее. Не реагирует она и на слишком мелкие предметы.
В поведении кольчатых червей еще преобладает избегание неблагоприятных внешних условий, как это имеет место у простейших. Но все же активный поиск положительных раздражителей занимает в поведении кольчецов уже заметное место, и это весьма характерно для высшего уровня элементарной сенсорной психики. Так же как и у простейших, в жизни кольчатых червей и других низших многоклеточных беспозвоночных большую роль играют кинезы и элементарные таксисы. Но наряду с ними здесь уже встречаются зачатки сложных форм инстинктивного поведения (особенно у некоторых многощетинковых червей, пиявок, а также улиток) и впервые появляются высшие таксисы, обеспечивающие значительно более точную и экономную ориентацию животного в пространстве, а тем самым и более полноценное использование пищевых ресурсов в окружающей среде. В результате возникли предпосылки для поднятия всей жизнедеятельности на более высокую ступень, что и имело место на стадии перцептивной психики.
Как было показано, у высших представителей рассматриваемой группы беспозвоночных впервые появляются зачатки конструктивной деятельности, агрессивного поведения, общения. Здесь опять подтверждается правило, что высшие формы поведения зарождаются уже на низших стадиях развития психической деятельности.
Давая общую оценку поведению низших многоклеточных беспозвоночных, следует отметить, что, очевидно, первично главная функция еще примитивной нервной системы состояла в координации внутренних процессов жизнедеятельности в связи со все большей специализацией клеток и новых образований - тканей, из которых строятся все органы и системы многоклеточного организма. «Внешние» же функции нервной системы определяются степенью внешней активности, которая у этих животных находится на еще невысоком уровне, зачастую не более высоком, чем у высших представителей простейших. Вместе с тем строение и функции рецепторов, как и «внешняя» деятельность нервной системы, значительно усложняются у животных, ведущих более активный образ жизни. Особенно это относится к свободноживущим, активно передвигающимся формам.
Говоря об обширной и пестрой по составу группе низших беспозвоночных, нужно, конечно, учесть, что поведение этих животных еще очень слабо изучено. Мы, например, еще почти ничего не знаем об онтогенезе поведения этих животных, о том, как формируется и развивается их поведение, да и совершенствуется ли оно вообще в процессе индивидуального развития, на что уже указывалось в части, посвященной онтогенезу поведения. Вполне возможно (если, конечно, исключить метаморфозные преобразования, личиночное поведение у низших многоклеточных и т.п.), что подобное онтогенетическое совершенствование у обсуждаемых животных не является существенным или даже вообще не происходит, ибо жесткость врожденных программ поведения, исключительная стереотипия форм реагирования являются у этих животных определяющей чертой всего их поведения.
Глава 2 ПЕРЦЕПТИВНАЯ ПСИХИКА
Перцептивная психика является высшей стадией развития психического отражения. Напомним, что эта стадия характеризуется, по Леонтьеву, изменением строения деятельности - выделением содержания деятельности, относящейся к условиям, в которых дан объект деятельности в среде (операции). По этой причине мы встречаемся здесь уже с подлинными навыками и восприятиями. Предметные компоненты среды отражаются уже как целостные единицы, в то время как при элементарной сенсорной психике имело место отражение лишь отдельных их свойств или суммы последних. Предметное восприятие обязательно предполагает определенную степень обобщения, появляются чувственные представления.
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
На этом уровне находятся прежде всего высшие беспозвоночные - головоногие моллюски и членистоногие (по меньшей мере высшие). Характеристику рассматриваемого уровня психического развития мы дадим на примере насекомых - центрального класса типа членистоногих. (Кроме насекомых к членистоногим относятся ракообразные, паукообразные, многоножки и некоторые мелкие группы.) Насекомые - самый многочисленный класс животных как по количеству видов, так и по количеству особей. Насекомые обитают повсюду на суше (во всех климатических зонах) - как на поверхности, так и в почве, во всех пресных водоемах, а также в воздухе, поднимаясь на высоту до двух километров.
Движения
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции членистоногих, особенно насекомых, было завоевание новой среды обитания - суши. Членистоногие - первые наземные животные в истории Земли. Переход на сушу был сопряжен с развитием особых органов передвижения - конечностей в виде сложных рычагов, состоящих из отдельных, соединенных суставами члеников, по которым членистоногие и получили свое название. Такие сложные рычаги (их аналогами являются конечности наземных позвоночных) наилучшим образом служат для передвижения по твердому субстрату путем отталкивания от него. Членистоногие в состоянии передвигаться по суше почти всеми возможными способами (ползание, ходьба, бег как по горизонтальным, так и по вертикальным поверхностям, прыгание, передвижение в толще грунта). В воде членистоногие плавают, ныряют, ползают и даже бегают по ее поверхности.
У насекомых, кроме того, в связи с переходом к жизни в воздушной среде впервые появился и совершенно новый способ передвижения - полет с помощью крыльев. Насекомые способны развить в полете очень большую скорость: у некоторых стрекоз она достигает 90 километров в час, у самца овода - даже 122 километра в час. Поистине поразительна частота взмахов крыльев у многих насекомых. Так, мотылек делает лишь 9 ударов в секунду, пчела же - 190, а комнатная муха - 330, но у некоторых мух и перепончатокрылых скорость вибрации крыльев достигает до тысячи ударов в секунду. Столь быстрые движения и соответственно частоты мышечных сокращений не наблюдаются ни у каких других животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями: мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей, мышцы внутренних органов.
Как видно, мускулатура насекомых отличается высокой степенью дифференцированности и специализированности. К тому же подавляющее большинство их мышц - поперечнополосатые. Количество отдельных мускулов исключительно велико - полторы тысячи и более. Внушительна и сила мышц: насекомые способны таскать предметы, иногда в сто раз превышающие их собственный вес. В целом, как отмечал Догель, у членистоногих мышечная система беспозвоночных достигает «апогея своего развития» как в морфологическом, так и в физиологическом отношении.
Центральная нервная система
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
В связи с отчетливо выраженной цефализацией в головном отделе слившиеся ганглии образовали мощный головной мозг в виде сложно устроенного надглоточного узла, в котором различают три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Передний отдел, протоцеребрум, развит сильнее остальных. Особое внимание заслуживают находящиеся в нем так называемые грибовидные (или стебельчатые) тела - ассоциативные и координирующие центры. В наибольшей степени они развиты у насекомых с особенно сложным поведением - у пчел, прежде всего рабочих особей, и муравьев. С протоцеребрумом связана и пара очень крупных и сложно устроенных зрительных долей, иннервирующих сложные глаза. Дейтоцеребрум (средний отдел головного мозга) иннервирует усики, тритоцеребрум иннервирует область рта и, кроме того, тесно связан с симпатической нервной системой. Вообще для насекомых характерна большая концентрация нервных элементов. Такая концентрация повышает возможности управления отдельными системами органов и поднимает общий морфофизиологический уровень организма как целостной единицы.
Нервная система, и в частности головной мозг, устроена у разных членистоногих, однако, весьма различно. Существенные различия обнаруживаются между низшими и высшими представителями даже в пределах классов, а иногда и отрядов. Так, например, отношение размеров мозга к объему туловища равняется у майского жука 1: 3290, у муравья 1: 280, а у пчелы 1: 174.
На очень высоком уровне развития находится нервная система головоногих моллюсков, в корне отличаясь от таковой других моллюсков. Особенно это относится к их весьма сложно устроенному головному мозгу. Обращает на себя внимание исключительная концентрация мозговых ганглиев: ганглии слились воедино и образовали общую околопищеводную массу. Основания зрительных нервов образуют громадные вздутия - оптические ганглии, которые своими размерами нередко превосходят сами церебральные узлы.
Нервы, отходящие от мозга к эффекторам, достигают очень большой мощности - до миллиметра и больше в диаметре, что в 5-10 раз превышает толщину нервов большинства позвоночных! Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А.А.Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, - все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств.
Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения - своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков - фасеток. Каждой фасетке соответствует самостоятельный глазок - омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7- 8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2-3 сантиметров. От объектов, расположенных дальше (уже начиная с 5-7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т.е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т.е. оптимального для зрения пчелы, расстояния.
Пластичность поведения
Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих животных еще небольшую роль. Ассоциативные связи формируются у них с трудом и лишь в ограниченных пределах. Результаты научения сохраняются недолго. У всех кольчатых червей встречается наиболее простая форма научения - привыкание, с которым мы уже встречались у простейших, у которых она является главной, если не единственной формой модификации врожденного, видотипичного поведения. У низших многоклеточных животных врожденные реакции на определенные раздражения также прекращаются после многократного повторения, если не последует адекватного подкрепления этих реакций. Так, например, дождевые черви перестают реагировать на повторное затенение, если оно остается без последствий.
Отмечается привыкание и в сфере пищевого поведения: если многощетинкового червя повторно «кормить» комочками бумаги, смоченными соком его обычной жертвы, он перестает их принимать. Если попеременно с такими комочками давать ему подлинные кусочки пищи, то он в конце концов научается различать их и будет отвергать лишь несъедобную бумагу. Подобные опыты ставились и на кишечнополостных (на полипах, находящихся, как мы уже знаем, на более низком уровне элементарной сенсорной психики). При этом полипы вели себя так же, как морские черви: уже после нескольких (до 5) опытов они отбрасывали несъедобные объекты еще до того, как подносили их к ротовому отверстию. Интересно, что нетренированные щупальца этого не делали, даже находясь рядом с тренированными.
Эти эксперименты интересны тем, что показывают способность низших беспозвоночных отличить по побочным физическим качествам съедобное от несъедобного (по вкусу предлагавшиеся объекты были одинаковыми), что подтверждает наличие истинного психического отражения уже на этом, низшем, уровне филогенетического развития. Ведь здесь производится опосредствованное действие, отличающееся тем, что свойство (или сочетание нескольких свойств), которым животное руководствуется при оценке пригодности объекта к пищевому употреблению, выступает как подлинный сигнал, а чувствительность червя или полипа играет роль посредника между организмом и компонентом среды, от которого непосредственно зависит существование животного.
Более сложное научение путем «проб и ошибок» и формирования новой индивидуальной двигательной реакции можно в элементарной форме обнаружить, начиная уже с плоских червей. Так, планарии, встречая на своем пути полоску наждачной бумаги, сперва останавливаются, но затем все же переползают через нее. Если же сочетать соприкосновение червя с наждачной бумагой с его сотрясением, то планарии перестают переползать через нее и тогда, когда не производится сотрясения. Правда, здесь, очевидно, еще нет подлинной ассоциации между двумя раздражителями (шероховатость и сотрясение); скорее всего, происходит суммация первоначально не очень сильного отрицательного раздражения (шероховатость) с дополнительным отрицательным раздражением (сотрясение) и что, вероятно, влечет за собой общее повышение возбудимости животного.
Все же можно выработать уже у планарии и более сложные реакции, когда один из раздражителей является биологически «нейтральным». В таких случаях можно говорить, очевидно, уже об элементарных процессах подлинного ассоциативного научения. Так, например, Л.Г.Воронин и Н.А.Тушмалова сумели выработать у планарии и кольчатых червей оборонительные и пищевые условные рефлексы. При этом были получены и данные, свидетельствующие об усложнении форм временных связей. Если по поводу плоских червей можно говорить лишь о примитивных неустойчивых условных рефлексах, то у многощетинковых червей обнаруживаются уже стабильные самовосстанавливающиеся после угасания условные рефлексы. Эти различия соответствуют глубоким морфологическим различиям в строении нервной системы этих групп животных и свидетельствуют о существенном прогрессе психической активности у полихет.
Пластичность поведения дождевых червей очень убедительно была показана еще в 1912 г. известным американским зоопсихологом Р.Иерксом. В проведенных им опытах черви должны были в Т-образном лабиринте выбирать определенную сторону, где находилось «гнездо» (в противоположной стороне червь получал электрический удар). Чтобы этому научить червей, потребовалось 120- 180 опытов. (Улитки осваивают такую задачу после 60 опытов и запоминают верное решение в течение 30 дней.) Впоследствии аналогичные опыты с дождевыми червями ставились Л.Геком, И.С.Робинсоном, Р.И.Войтусяком (рис. 34) и другими исследователями. Удавалось также переучивать червей, поменяв «гнездо» и электроды местами. Характерно, что результат научения не меняется и после удаления передних сегментов. Более того, даже лишенный с самого начала этих сегментов, червь оказывается способным научиться правильно ориентироваться в лабиринте.
Подобные факты побудили в свое время В.А. Вагнера говорить о «сегментарной психологии» кольчатых червей и других «членистых», имея при этом в виду, что ганглии каждого сегмента в большой степени обеспечивают автономное выполнение элементарных психических функций. Обезглавленные кольчецы, писал Вагнер, «не теряют способности к спонтанным движениям, за исключением тех лишь, которые стоят в прямой зависимости и связи с органами чувств головы… Обезглавленные черви удерживают свои инстинктивные действия, даже те, в которых голова принимает прямое участие».*
* Вагнер В.А.Сегментарная психология. Новые идеи в биологии. Сборник № 6. Биопсихология. Спб., 1914. С. 123.
Рис. 34. Научение у дождевого червя (опыты Войтусяка)
Все это свидетельствует о том, что на уровне даже высших червей цефализация еще не достигла такого развития, чтобы оказать решающее влияние на все поведенческие акты животного. Все же общая направляющая роль головного мозга достаточно велика уже и на этом уровне. Это проявляется, например, в том, что если разрезать пополам дождевого червя, передняя половина будет закапываться в почву, производя вполне координированные движения, задняя же будет лишь беспорядочно извиваться.
Еще лучше, чем у малощетинковых червей, как уже отмечалось, вырабатываются ассоциативные связи у полихет. В опытах удавалось, например, переделать отрицательную реакцию на освещение в положительную путем сочетания освещения с подкармливанием. В итоге черви стали при освещении выползать из своих домиков и без пищевого подкрепления. Время реакции (между световым раздражением и ответным движением) уменьшилось при этом с 28,9 до 3,5 секунды (опыты М.Купленда).
Обстоятельно изучал способность к научению полихет С.М.Эванс. Применяя наказания электрическим током, этот исследователь довольно легко выработал у подопытных животных отрицательную двигательную реакцию (отказ от вползания в узкий темный коридор). В других опытах червям приходилось проползать Т-образный лабиринт. При правильном повороте они могли в течение 5 минут пребывать в затемненной камере. Особенно хорошо обучались полихеты Nereis virens, которым понадобилось менее 100 сочетаний для почти безошибочного решения задачи и ее прочного запоминания; спустя 48 часов они безошибочно воспроизводили правильный путь через лабиринт (рис. 35).
Рис. 35. Научение у Nereis virens (опытЭванса). Вверху даны схемы Т-образных лабиринтов, в которых производились опыты. Внизу - кривая научения (по оси ординат-число ошибок, по оси абсцисс - число опытов)
Общая характеристика высшего уровня элементарной сенсорной психики
Как уже отмечалось, наиболее низкоорганизованные формы многоклеточных беспозвоночных стоят на том же уровне психического развития, что и высшие представители простейших. Но то, что здесь не обнаруживаются существенные различия в поведении, несмотря на глубокие различия в строении, не должно нас удивлять, ибо, как уже говорилось, простейшие олицетворяют собой совершенно особую, рано отклонившуюся филогенетическую ветвь, которая до известных пределов развивалась параллельно ветви низших многоклеточных животных.
Что же касается поведения кольчатых червей, то оно вполне отвечает стадии элементарной сенсорной психики, ибо слагается из движений, ориентированных лишь по отдельным свойствам предметов и явлений (или их сочетаниям), причем это те свойства, которые, по Леонтьеву, оповещают о появлении жизненно важных условий среды, от которых зависит осуществление основных биологических функций животных. Эта ориентация осуществляется, таким образом, на основе одних лишь ощущений. Перцепция, способность к предметному восприятию, еще отсутствует. Не исключено, правда, что у некоторых наземных улиток, как и у упомянутых выше свободно плавающих хищных моллюсков и полихет, уже намечаются зачатки этой способности. Так, виноградная улитка обходит преграду еще до прикосновения к ней, ползет вдоль нее, но только если преграда не слишком велика; если же изображение преграды занимает всю сетчатку, улитка наталкивается на нее. Не реагирует она и на слишком мелкие предметы.
В поведении кольчатых червей еще преобладает избегание неблагоприятных внешних условий, как это имеет место у простейших. Но все же активный поиск положительных раздражителей занимает в поведении кольчецов уже заметное место, и это весьма характерно для высшего уровня элементарной сенсорной психики. Так же как и у простейших, в жизни кольчатых червей и других низших многоклеточных беспозвоночных большую роль играют кинезы и элементарные таксисы. Но наряду с ними здесь уже встречаются зачатки сложных форм инстинктивного поведения (особенно у некоторых многощетинковых червей, пиявок, а также улиток) и впервые появляются высшие таксисы, обеспечивающие значительно более точную и экономную ориентацию животного в пространстве, а тем самым и более полноценное использование пищевых ресурсов в окружающей среде. В результате возникли предпосылки для поднятия всей жизнедеятельности на более высокую ступень, что и имело место на стадии перцептивной психики.
Как было показано, у высших представителей рассматриваемой группы беспозвоночных впервые появляются зачатки конструктивной деятельности, агрессивного поведения, общения. Здесь опять подтверждается правило, что высшие формы поведения зарождаются уже на низших стадиях развития психической деятельности.
Давая общую оценку поведению низших многоклеточных беспозвоночных, следует отметить, что, очевидно, первично главная функция еще примитивной нервной системы состояла в координации внутренних процессов жизнедеятельности в связи со все большей специализацией клеток и новых образований - тканей, из которых строятся все органы и системы многоклеточного организма. «Внешние» же функции нервной системы определяются степенью внешней активности, которая у этих животных находится на еще невысоком уровне, зачастую не более высоком, чем у высших представителей простейших. Вместе с тем строение и функции рецепторов, как и «внешняя» деятельность нервной системы, значительно усложняются у животных, ведущих более активный образ жизни. Особенно это относится к свободноживущим, активно передвигающимся формам.
Говоря об обширной и пестрой по составу группе низших беспозвоночных, нужно, конечно, учесть, что поведение этих животных еще очень слабо изучено. Мы, например, еще почти ничего не знаем об онтогенезе поведения этих животных, о том, как формируется и развивается их поведение, да и совершенствуется ли оно вообще в процессе индивидуального развития, на что уже указывалось в части, посвященной онтогенезу поведения. Вполне возможно (если, конечно, исключить метаморфозные преобразования, личиночное поведение у низших многоклеточных и т.п.), что подобное онтогенетическое совершенствование у обсуждаемых животных не является существенным или даже вообще не происходит, ибо жесткость врожденных программ поведения, исключительная стереотипия форм реагирования являются у этих животных определяющей чертой всего их поведения.
Глава 2 ПЕРЦЕПТИВНАЯ ПСИХИКА
Низший уровень развития перцептивной психики
Перцептивная психика является высшей стадией развития психического отражения. Напомним, что эта стадия характеризуется, по Леонтьеву, изменением строения деятельности - выделением содержания деятельности, относящейся к условиям, в которых дан объект деятельности в среде (операции). По этой причине мы встречаемся здесь уже с подлинными навыками и восприятиями. Предметные компоненты среды отражаются уже как целостные единицы, в то время как при элементарной сенсорной психике имело место отражение лишь отдельных их свойств или суммы последних. Предметное восприятие обязательно предполагает определенную степень обобщения, появляются чувственные представления.
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
На этом уровне находятся прежде всего высшие беспозвоночные - головоногие моллюски и членистоногие (по меньшей мере высшие). Характеристику рассматриваемого уровня психического развития мы дадим на примере насекомых - центрального класса типа членистоногих. (Кроме насекомых к членистоногим относятся ракообразные, паукообразные, многоножки и некоторые мелкие группы.) Насекомые - самый многочисленный класс животных как по количеству видов, так и по количеству особей. Насекомые обитают повсюду на суше (во всех климатических зонах) - как на поверхности, так и в почве, во всех пресных водоемах, а также в воздухе, поднимаясь на высоту до двух километров.
Движения
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции членистоногих, особенно насекомых, было завоевание новой среды обитания - суши. Членистоногие - первые наземные животные в истории Земли. Переход на сушу был сопряжен с развитием особых органов передвижения - конечностей в виде сложных рычагов, состоящих из отдельных, соединенных суставами члеников, по которым членистоногие и получили свое название. Такие сложные рычаги (их аналогами являются конечности наземных позвоночных) наилучшим образом служат для передвижения по твердому субстрату путем отталкивания от него. Членистоногие в состоянии передвигаться по суше почти всеми возможными способами (ползание, ходьба, бег как по горизонтальным, так и по вертикальным поверхностям, прыгание, передвижение в толще грунта). В воде членистоногие плавают, ныряют, ползают и даже бегают по ее поверхности.
У насекомых, кроме того, в связи с переходом к жизни в воздушной среде впервые появился и совершенно новый способ передвижения - полет с помощью крыльев. Насекомые способны развить в полете очень большую скорость: у некоторых стрекоз она достигает 90 километров в час, у самца овода - даже 122 километра в час. Поистине поразительна частота взмахов крыльев у многих насекомых. Так, мотылек делает лишь 9 ударов в секунду, пчела же - 190, а комнатная муха - 330, но у некоторых мух и перепончатокрылых скорость вибрации крыльев достигает до тысячи ударов в секунду. Столь быстрые движения и соответственно частоты мышечных сокращений не наблюдаются ни у каких других животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями: мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей, мышцы внутренних органов.
Как видно, мускулатура насекомых отличается высокой степенью дифференцированности и специализированности. К тому же подавляющее большинство их мышц - поперечнополосатые. Количество отдельных мускулов исключительно велико - полторы тысячи и более. Внушительна и сила мышц: насекомые способны таскать предметы, иногда в сто раз превышающие их собственный вес. В целом, как отмечал Догель, у членистоногих мышечная система беспозвоночных достигает «апогея своего развития» как в морфологическом, так и в физиологическом отношении.
Центральная нервная система
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
В связи с отчетливо выраженной цефализацией в головном отделе слившиеся ганглии образовали мощный головной мозг в виде сложно устроенного надглоточного узла, в котором различают три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Передний отдел, протоцеребрум, развит сильнее остальных. Особое внимание заслуживают находящиеся в нем так называемые грибовидные (или стебельчатые) тела - ассоциативные и координирующие центры. В наибольшей степени они развиты у насекомых с особенно сложным поведением - у пчел, прежде всего рабочих особей, и муравьев. С протоцеребрумом связана и пара очень крупных и сложно устроенных зрительных долей, иннервирующих сложные глаза. Дейтоцеребрум (средний отдел головного мозга) иннервирует усики, тритоцеребрум иннервирует область рта и, кроме того, тесно связан с симпатической нервной системой. Вообще для насекомых характерна большая концентрация нервных элементов. Такая концентрация повышает возможности управления отдельными системами органов и поднимает общий морфофизиологический уровень организма как целостной единицы.
Нервная система, и в частности головной мозг, устроена у разных членистоногих, однако, весьма различно. Существенные различия обнаруживаются между низшими и высшими представителями даже в пределах классов, а иногда и отрядов. Так, например, отношение размеров мозга к объему туловища равняется у майского жука 1: 3290, у муравья 1: 280, а у пчелы 1: 174.
На очень высоком уровне развития находится нервная система головоногих моллюсков, в корне отличаясь от таковой других моллюсков. Особенно это относится к их весьма сложно устроенному головному мозгу. Обращает на себя внимание исключительная концентрация мозговых ганглиев: ганглии слились воедино и образовали общую околопищеводную массу. Основания зрительных нервов образуют громадные вздутия - оптические ганглии, которые своими размерами нередко превосходят сами церебральные узлы.
Нервы, отходящие от мозга к эффекторам, достигают очень большой мощности - до миллиметра и больше в диаметре, что в 5-10 раз превышает толщину нервов большинства позвоночных! Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А.А.Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, - все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств.
Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения - своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков - фасеток. Каждой фасетке соответствует самостоятельный глазок - омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7- 8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2-3 сантиметров. От объектов, расположенных дальше (уже начиная с 5-7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т.е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т.е. оптимального для зрения пчелы, расстояния.