Страница:
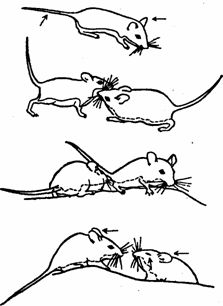
Рис. 43. Оптико-ольфакторное общение у грызунов (род Peromyscus). Стрелками обозначены места взаимного обнюхивания (по Эйзенбергу)
Для биологически адекватного реагирования на все эти звуковые сигналы существенной является видоспецифическая настройка слуховой системы на определенную часть спектра. Как показал советский исследователь биоакустики птиц В.Д.Ильичев, важнейшую роль играют в этой настройке периферические отделы слухового анализатора, выполняющие роль видотипических биологических фильтров. С их помощью осуществляется и специальная «подстройка» слуховой системы на особенно важные для особи звуки. Таким образом, здесь четко выступает характерная для всего инстинктивного поведения взаимообусловленность и единство врожденных функциональных (физиологических и этологических) и морфологических элементов.
Зоосемантика звуковых сигналов птиц охватывает практически все сферы их жизнедеятельности. Эти сигналы служат опознаванию особи (в частности, ее видовой принадлежности), оповещению других особей о физиологическом состоянии экспедиента (сюда относится, например, «попрошайничество») или об изменениях в окружающей среде (появление врага или, наоборот, кормового объекта и т.п.). При этом богатые зоопрагматические средства птицы позволяют экспедиенту передавать перцепиенту весьма детализованную информацию. Так, грач, подлетающий с кормом к гнезду, оповещает об этом насиживающую самку криком, могущим иметь четыре разных оттенка, на которые самка также отвечает разными звуками. Эти оттенки голосовых реакций птиц подчас настолько тонки, что не воспринимаются человеческим ухом.
Очень большое значение имеют не только четкие межвидовые различия акустического общения, но и индивидуальные, по которым особи узнают друг друга (самец - самку, птенцы - родителей и друг друга, члены стаи - друг друга и т.п.). Интересно, например, что, по данным В.Г.Торпа, в ряде случаев совместно поющие партнеры откликаются только на голос друг друга, но не реагируют на пение других особей. Особенно индивидуально различными являются территориальные крики птиц, оповещающих о занятости участка.
Большого разнообразия звуки достигают и у таких птиц, как куриные. Это обусловлено жизнью диких кур в труднопросматриваемых густых кустарниковых зарослях, где условия для оптического общения особенно неблагоприятны. Наиболее громкие и «впечатляющие» звуки куриных птиц - это крики петухов. Их слышимость (для человеческого уха) достигает 2 километров. В отличие, например, от звуков, с помощью которых петух подзывает кур к найденному им корму, кукареканье производится однократно, но зато дольше и на более высоких частотах. Это относится и к звуковым сигналам тревоги (например, при обнаружении врага), испуга или угрозы. Чем выше тональность, громкость и длительность этих сигналов, тем сильнее их эффект. Свое «кукареку» петух кричит на самых «высоких нотах», но всегда одинаковым образом: отклонения не превышают полтона.
Кукареканье - это своего рода вызов другим петухам, и те отвечают ответными криками, правда, лишь в том случае, если соперники - петухи одного «ранга». Если же в пределах видимости закричит петух, занимающий подчиненное положение, то господствующий попросту атакует его, «не удостаивая» ответным криком. Когда же перекликаются петухи, разделенные большим расстоянием и не видящие друг друга, то это типичные территориальные крики, т.е. акустическая маркировка местности, оповещение о занятости участков стайками кур - петухом и его «гаремом». Большое же биологическое значение крика заключается в том, что он предотвращает или хотя бы уменьшает частоту петушиных боев, а вместе с тем обеспечивает захват, освоение и защиту участков, а в конечном итоге расселение вида, но безвредным путем, ибо дело ограничивается одним запугиванием. Петух-соперник заранее предупреждается о том, что данный участок уже занят и ему придется искать другое пригодное для поселения место. И только в том случае, если все такие места уже заняты, начнутся бои за овладение участками. В этом случае победитель - животное более сильное не только в физическом, но и в психическом отношении - станет продолжателем рода, и это, конечно, тоже выгодно для процветания вида, для его прогрессивной эволюции. Аналогичную роль играет у других птиц пение. У домашних же кур крестьянский двор и прилегающие места остаются для петуха его участком, о занятии которого он непрестанно напоминает криком, хотя это поведение уже потеряло свое биологическое значение, поскольку человек обеспечивает все потребности домашних животных и управляет эволюцией вида. Надо думать, что чрезвычайная близость расположения участков домашних петухов заставляет их интенсивнее кукарекать, чем это имело бы место в дикой природе, так как в селе популяция кур «сверхплотная».
Мы не останавливаемся на звуковом общении млекопитающих, так как у них господствуют те же закономерности, что и у птиц.
Ригидность и пластичность в поведении высших позвоночных
Как уже указывалось, вопреки еще распространенному мнению инстинктивное поведение не теряет своей значимости в процессе эволюции, так как оно принципиально не может замещаться научением. Подчеркнем еще раз, что инстинктивное поведение является видовым поведением, научение - индивидуальным, и поэтому ничем не оправдано противопоставление этих двух основных категорий психической деятельности в качестве якобы различных генетических ступеней. На самом деле, как мы могли убедиться, прогресс инстинктивного поведения, в частности, в ряду позвоночных неразрывно связан с прогрессом индивидуально-изменчивого поведения, поэтому позвоночные с высокоразвитыми формами научения обладают и не менее развитыми сложными формами инстинктивного поведения.
Вместе с тем темпы эволюционных преобразований и специфическая роль в эволюции у инстинктивного поведения и научения различны. Главным образом, как показал Северцов, это относится к высшим позвоночным, особенно млекопитающим, у которых психика приобретает значение решающего фактора эволюции благодаря сильному развитию процессов научения, в том числе в высших их проявлениях - интеллектуальных действиях. Но при этом сохраняется в полной мере значение инстинктивной основы поведения, равно как медленно совершающихся изменений инстинктивного поведения. Приведенные примеры в достаточной мере показывают большое разнообразие истинных инстинктивных движений у высших позвоночных, вполне видотипичных, генетически фиксированных и достигающих в ряде случаев большой сложности. Конечно, и у высших позвоночных инстинктивные компоненты служат прежде всего для пространственно-временной ориентации наиболее жизненно важных поведенческих актов. Пространственная ориентация осуществляется и здесь на основе таксисов: тропо-, тело- и менотаксисов, т.е. типичных врожденных элементов поведения, к которым, однако, прибавляются мнемотаксисы, характеризующиеся индивидуальным запоминанием ориентиров. В последнем случае мы уже имеем дело с накоплением индивидуального опыта, и поскольку именно мнемотаксисы играют в жизни высших позвоночных особенно большую роль, то, следовательно, врожденные способы ориентации существенно обогащаются здесь индивидуально приобретаемыми способами.
В этой связи следует отметить, что, как и у других животных, биологическая адекватность реагирования на компоненты окружающей среды обеспечивается и у высших позвоночных именно инстинктивными элементами поведения. Биологически значимые объекты встречаются в окружающей животное среде в весьма разнообразных и, главное, постоянно меняющихся видах. Мы уже знаем, что их потребление или избегание, т.е. адекватное реагирование на биологические ситуации, возможно лишь в том случае, если животное руководствуется некими постоянными признаками этих объектов и ситуации. Именно это и происходит на генетически фиксированной, врожденной основе, когда животное реагирует на ключевые раздражители. К высшим позвоночным это относится точно так же, как к нижестоящим животным. Но у первых реакции на ключевые раздражители в очень большой степени конкретизируются индивидуальным опытом, дополнительно ориентируются хорошо развитым предметным восприятием. Тем самым инстинктивные действия приобретают и известное познавательное значение для животного, ибо помогают ему при ознакомлении с окружающей действительностью.
Особенно высокого уровня развития инстинктивное поведение достигает у высших позвоночных в ритуализованном общении животных друг с другом. Именно в сфере общения инстинктивные формы поведения достигают наибольшей стереотипности. Совершенно ясно, что иначе, без жестко фиксированных зоопрагматических средств, не может быть взаимопонимания между животными, т.е. не может быть подлинной передачи информации. Вместе с тем именно полноценное общение является необходимым условием для высшей интеграции в области поведения - интеграции поведения отдельных особей и целых сообществ.
Однако даже в сфере общения наличие и важное значение благоприобретаемых компонентов не вызывает сомнения. Так, у многих птиц птенцы не смогут петь видотипичным образом, если своевременно не слышали пения своих родителей. Несомненна роль научения и в образовании индивидуальных особенностей звукового общения, а также в многочисленных случаях акустического подражания у взрослых птиц. В последнем случае оказалось, что заимствованные звуки могут служить для общения наряду с собственными. На этой основе развивается межвидовое общение птиц, т.е. передача информации между особями разных видов. Здесь индивидуально приобретенные коммуникативные компоненты с четким сигнальным значением уже не только модифицируют и обогащают собственно инстинктивные компоненты, но и выступают во вполне самостоятельной роли.
Индивидуально приобретаемые элементы общения представляют особый «интерес, так как показывают возможность выхода за пределы закрытых коммуникативных систем животных. Особенно это относится к общению животных с человеком, в частности при дрессировке служебных собак и т.п. Вероятно, расширенное общение с животными сыграло в свое время немаловажную роль и при одомашнивании диких видов. Все это, разумеется, оказалось возможным лишь на высшем уровне перцептивной психики, т.е. у высших позвоночных. Если же иметь в виду предысторию антропогенеза, о чем пойдет речь ниже, то нельзя не признать, что способность высших позвоночных к расширению своих коммуникативных возможностей путем научения должна была стать важной предпосылкой зарождения человеческих форм общения.
Сказанное здесь о роли благоприобретаемых компонентов в инстинктивном поведении высших позвоночных вполне относится и к другим сферам поведения. Напомним, например, что у зубров и бизонов видотипичные признаки поведения появляются лишь на определенном этапе онтогенеза. Хотя здесь, несомненно, имеет место созревание некоторых видотипичных двигательных компонентов вне зависимости от частных внешних условий, все же и в этих процессах участвуют элементы научения. В остальном ограничимся здесь отсылкой к тому, что раньше говорилось по этому поводу об онтогенезе поведения.
Нет также надобности еще раз говорить о том, что в разных поведенческих актах удельный вес врожденных и благоприобретаемых компонентов различен, что и в навыках позвоночных содержатся качественно-гетерогенные элементы. Напомним лишь, что, не говоря уже о том, что навыки формируются на основе безусловнорефлекторных процессов, в их состав всегда входят консервативные двигательные компоненты, придающие навыкам в некоторых отношениях сходство с инстинктивными действиями. Более того, именно формирование таких консервативных компонентов составляет во многих случаях конечный итог и биологическую сущность образования навыка. Это. относится к двигательным стереотипам, автоматизмам, возникающим как результат закрепления навыка в ходе тренировки. Столь примитивные навыки встречаются не только у низших позвоночных. Наоборот, заученные автоматизированные действия играют немаловажную роль и в жизни высших млекопитающих, включая обезьян, а также человека (например, техника письма, повседневные «бытовые» движения и т.п.). У рыб же такими элементарными навыками, очевидно, ограничиваются их способности к научению. С этим связан и тот факт, что у рыб крайне трудно, если вообще возможно переделать положительное значение раздражителя на отрицательное и наоборот. Причиной тому является, в частности, недостаточное развитие ассоциативных функций головного мозга низших позвоночных. Как показал советский физиолог Б.Ф.Сергеев, только на уровне костистых рыб и земноводных появляется способность к образованию временных связей между всеми анализаторными системами и функциями организма, но еще отсутствуют внутри- и межанализаторные ассоциативные временные связи. Анализаторные системы еще разобщены, локализованы в различных отделах мозга.
Только у птиц и млекопитающих замыкательная функция головного мозга получает свое полное развитие, поэтому сложные навыки, в которых решающее значение имеют лабильные, гибкие компоненты, встречаются только у высших позвоночных. Они и определяют далеко идущую изменчивость, пластичность всего поведения животного, чему придавал столь большое адаптационное значение А.Н.Северцов. И действительно, сложные пластичные навыки в полной мере выполняют функцию быстрого приспособления организма к быстротечным изменениям среды. Пластичность навыков высшего порядка дополняет ригидность навыков низшего порядка и инстинктивных действий.
Эта пластичность проявляется в возможности быстрой перестройки навыка, в частности при превращении положительного или отрицательного раздражителя в противоположный. Другая важная особенность - возможность переноса навыка в новые условия, другими словами, адекватное использование накопленного чувственного и моторного опыта при существенных изменениях условий среды. Обеспечиваются эти возможности сильным развитием пластичности в сенсорной сфере, способностью к широким чувственным обобщениям, о чем уже шла речь выше.
Поясним сказанное на нескольких примерах. В первом эксперименте крыса получает приманку в результате ряда последовательных сложных инструментальных действий: она взбирается по лестнице на площадку, поднимает там эту лестницу с помощью перекинутой через блок бечевки, затем поднимается по лестнице на следующую площадку и там получает пищевое подкрепление. К этим фазам можно было бы добавить еще несколько - суть дела от этого не меняется: мы имеем здесь сложную операцию, в которой последовательность отдельных действий животного предопределена и строго фиксирована условиями задачи, т.е. теми компонентами среды (преградами), на которые по необходимости направлена активность животного.
В другом примере при относительно простой структуре операции выделяется способность к далеко идущему переносу операции в измененных условиях, что также весьма характерно для сложных навыков. В данном случае крыса, научившаяся находить кратчайший путь к «цели» в лабиринте, оказалась без какого-либо нового обучения способной к этому и после того, как лабиринт был поставлен вертикально. Поскольку животное при этом руководствуется и новыми ощущениями (с вестибулярного аппарата), то здесь с очевидностью имеют место и явления переноса в сенсорной сфере.
В третьем эксперименте крыса, освоившая сухопутный лабиринт, тут же с неменьшим успехом проплыла этот лабиринт после того, как он был заполнен водой. Поскольку при плавании производятся иные движения, чем при ходьбе, мы можем здесь констатировать далеко идущий перенос в моторной сфере при сохранении прежней ориентации в пространстве.
Даже в первом примере, а тем более в остальных, невозможно расценивать поведение подопытного животного как простую цепь механически усвоенных движений, ибо в таком случае совершенно исключалась бы возможность переноса операции и сохранения навыка в измененных условиях. Сложные навыки представляют собой исключительно динамические моторнорецепторные системы, обеспечивающие на основе высокоразвитой ориентировочной деятельности выработку весьма пластичных двигательных программ. Процесс ориентировки сливается здесь с двигательной активностью, а нахождение верного решения задачи формируется в ходе этой активности на основе высокоразвитого чувственного обобщения.
Отмеченными качествами сложные навыки выделяются среди других видов научения, и именно благодаря им сложные навыки стали предпосылками и основой развития высших форм психической деятельности животных - интеллектуальных действий.
Проблема интеллекта животных
Предпосылки и элементы интеллектуального поведения животных
Интеллектуальное поведение является вершиной психического развития животных. Однако, говоря об интеллекте, «уме» животных, их мышлении необходимо прежде всего отметить, что чрезвычайно трудно точно указать, по поводу каких животных можно говорить об интеллектуальном поведении, а по поводу каких - нет. Очевидно, речь может идти лишь о высших позвоночных, но явно не только о приматах, как это до недавнего времени принималось. Вместе с тем интеллектуальное поведение животных является не чем-то обособленным, из ряда вон выходящим, а лишь одним из проявлений единой психической деятельности с ее врожденными и благоприобретаемыми аспектами. Интеллектуальное поведение не только теснейшим образом связано с разными формами инстинктивного поведения и научения, но и само складывается (на врожденной основе) из индивидуально-изменчивых компонентов поведения. Оно является высшим итогом и проявлением индивидуального накопления опыта, особой категорией научения с присущими ей качественными особенностями. Поэтому интеллектуальное поведение дает наибольший приспособительный эффект, на что и обратил особое внимание А.Н.Северцов, показав решающее значение высших психических способностей для выживания особей и продолжения рода при резких, быстро протекающих изменениях в среде обитания.
Предпосылкой и основой развития интеллекта животных - во всяком случае в направлении, ведущем к человеческому сознанию - является манипулирование, причем прежде всего с биологически «нейтральными» объектами. Особенно, как уже было показано, это относится к обезьянам, для которых манипулирование служит источником наиболее полных сведений о свойствах и структуре предметных компонентов среды, ибо в ходе манипулирования происходит наиболее глубокое и всестороннее ознакомление с новыми предметами или новыми свойствами уже знакомых животному объектов. В ходе манипулирования, особенно при выполнении сложных манипуляций, происходит обобщение опыта деятельности животного, формируются обобщенные знания о предметных компонентах окружающей среды, и именно этот обобщенный двигательно-сенсорный опыт составляет главнейшую основу интеллекта обезьян.
О манипулировании человекообразных обезьян «биологически индифферентными» предметами Павлов говорил: «Это же - самая настойчивая любознательность. Так что нелепое утверждение, будто у животных ее нет, нет в зачатке того, что есть у нас и что в конечном счете создало науку, - не отвечает действительности». [34]В качестве примера Павлов сослался на наблюдавшееся им у шимпанзе манипулирование предметами, в частности коробкой, в которой нет «никаких апельсинов, ни яблок». Тем не менее обезьяна «долгое время возится… над решением механических задач, которое не обещает ей никаких выгод, никакого материального удовлетворения».
Эта, по Павлову, «чистейшая, бескорыстная любознательность» и заставляет обезьяну изучать объект манипулирования в ходе активного воздействия на него. При этом одновременно и во взаимодействии друг с другом включаются в познавательную деятельность животного разные сенсорные и эффекторные системы. Ведь манипулирующая обезьяна почти непрерывно следит за движениями своих рук; под пристальным зрительным контролем производятся самые разнообразные действия как без разрушения целостности объекта: поворачивание в разные стороны, облизывание, поглаживание, придавливание, перекатывание и т.п., так и деструктивного порядка: разламывание, разрывание, вычленение отдельных деталей и т.д. (рис. 24).
Особую познавательную ценность представляют деструктивные действия, так как они позволяют получить сведения о внутренней структуре предметов. При манипулировании животное получает информацию одновременно по ряду сенсорных каналов, но преобладающее значение имеет у обезьян сочетание кожно-мышечной чувствительности рук со зрительными ощущениями. Кроме того, в обследовании объекта манипулирования участвуют также обоняние, вкус, тактильная чувствительность околоротовых вибрисс, иногда слух и т.д. Эти виды чувствительности сочетаются с кожно-мышечной чувствительностью эффекторов (ротового аппарата, передних конечностей) кроме обезьян и у других млекопитающих, когда те манипулируют предметами. В итоге животные получают комплексную информацию об объекте как едином целом и обладающем разнокачественньми свойствами. Именно в этом и заключается значение манипулирования как основы интеллектуального поведения.
Необходимо, однако, подчеркнуть, что первостепенное значение имеют для интеллектуального поведения зрительные восприятия и особенно зрительные обобщения, о которых уже раньше шла речь. Насколько развита способность к формированию обобщенных зрительных образов даже у крыс - показывает следующий эксперимент, в котором крысы с успехом решали очень трудную задачу: животному надо выбрать из трех предъявленных фигур (вертикальные и горизонтальные полосы) одну несходную по сравнению с двумя другими. Местонахождение и рисунок такой фигуры постоянно меняются, следовательно, это будут то вертикальные, то горизонтальные полосы, расположенные то слева, то справа, то посередине (в неправильной последовательности). Таким образом, подопытное животное могло ориентироваться лишь по одному, крайне обобщенному признаку - несхожести одного рисунка по сравнению с остальными. Мы имеем здесь, таким образом, дело со зрительным обобщением, близким к абстрагированию, свойственному мыслительным процессам.
С другим элементом интеллектуального поведения, на этот раз в двигательной сфере, мы встречались при описании опытов с «проблемными ящиками». И здесь мы имеем дело со сложными многофазовыми навыками, поскольку у высших млекопитающих, например енотов, можно сравнительно легко добиться решения задач, при которых животному приходится в определенной последовательности открывать совокупность разнообразных запирающих приспособлений. Как и при описанном опыте с подтягиванием крысой лестницы, енот может решить такую задачу лишь при соблюдении строго определенной последовательности действий. Но разница заключается в том, что в отличие от крысы еноту приходится самому находить эту последовательность, и это поднимает его деятельность, безусловно, на более высокий уровень. Правда, по некоторым данным и крысы способны на это.
Необходимо, однако, подчеркнуть, что даже высшие позвоночные решают инструментальные задачи труднее, чем локомоторные. Л. Кардош отметил в этой связи, что в психической деятельности животных преобладает познавание пространственных отношений (см. ч. I, гл. 3), постигаемых ими с помощью локомоторных действий. У обезьян же, особенно человекообразных, локомоторное познавание пространственных отношений теряет свою доминирующую роль за счет сильного развития манипуляционных действий. Однако только человек может полностью освободиться от направляющего воздействия пространственных отношений, если этого требует познание временно-причинных связей.
Решение многофазовых инструментальных задач у обезьян изучалось рядом исследователей, в частности Н. Н. Ладыгиной-Котс. В своей монографии «Приспособительные моторные навыки макака в условиях эксперимента» она подвела итог многочисленным опытам, в которых использовались весьма разнообразные сочетания запирающих механизмов. Эти эксперименты показали, что низшая обезьяна (макак-резус) способна научиться отмыкать большие серии запирающих механизмов, хотя лучше справляется с одиночными установками. Характерными явились при этом многочисленные и многообразные нащупывающие движения рук, «экспериментирование». Ввиду ее большой торопливости наиболее трудными оказались для обезьяны те приспособления, которые не отмыкались легкими быстрыми движениями. Легче всего производились такие движения, как вытягивание, отведение, притягивание, опускание и др., труднее всего - отодвигание и вращение, вывертывание. Вообще в поисках пунктов задержки и пути преодоления препятствий преобладающая роль принадлежала кинестетическим, а не зрительным восприятиям. Интересно, что по многим из этих особенностей, как показали более поздние исследования, действия низших обезьян в опытах с запирающими механизмами напоминают таковые енотов.
Интеллектуальное поведение является вершиной психического развития животных. Однако, говоря об интеллекте, «уме» животных, их мышлении необходимо прежде всего отметить, что чрезвычайно трудно точно указать, по поводу каких животных можно говорить об интеллектуальном поведении, а по поводу каких - нет. Очевидно, речь может идти лишь о высших позвоночных, но явно не только о приматах, как это до недавнего времени принималось. Вместе с тем интеллектуальное поведение животных является не чем-то обособленным, из ряда вон выходящим, а лишь одним из проявлений единой психической деятельности с ее врожденными и благоприобретаемыми аспектами. Интеллектуальное поведение не только теснейшим образом связано с разными формами инстинктивного поведения и научения, но и само складывается (на врожденной основе) из индивидуально-изменчивых компонентов поведения. Оно является высшим итогом и проявлением индивидуального накопления опыта, особой категорией научения с присущими ей качественными особенностями. Поэтому интеллектуальное поведение дает наибольший приспособительный эффект, на что и обратил особое внимание А.Н.Северцов, показав решающее значение высших психических способностей для выживания особей и продолжения рода при резких, быстро протекающих изменениях в среде обитания.
Предпосылкой и основой развития интеллекта животных - во всяком случае в направлении, ведущем к человеческому сознанию - является манипулирование, причем прежде всего с биологически «нейтральными» объектами. Особенно, как уже было показано, это относится к обезьянам, для которых манипулирование служит источником наиболее полных сведений о свойствах и структуре предметных компонентов среды, ибо в ходе манипулирования происходит наиболее глубокое и всестороннее ознакомление с новыми предметами или новыми свойствами уже знакомых животному объектов. В ходе манипулирования, особенно при выполнении сложных манипуляций, происходит обобщение опыта деятельности животного, формируются обобщенные знания о предметных компонентах окружающей среды, и именно этот обобщенный двигательно-сенсорный опыт составляет главнейшую основу интеллекта обезьян.
О манипулировании человекообразных обезьян «биологически индифферентными» предметами Павлов говорил: «Это же - самая настойчивая любознательность. Так что нелепое утверждение, будто у животных ее нет, нет в зачатке того, что есть у нас и что в конечном счете создало науку, - не отвечает действительности». [34]В качестве примера Павлов сослался на наблюдавшееся им у шимпанзе манипулирование предметами, в частности коробкой, в которой нет «никаких апельсинов, ни яблок». Тем не менее обезьяна «долгое время возится… над решением механических задач, которое не обещает ей никаких выгод, никакого материального удовлетворения».
Эта, по Павлову, «чистейшая, бескорыстная любознательность» и заставляет обезьяну изучать объект манипулирования в ходе активного воздействия на него. При этом одновременно и во взаимодействии друг с другом включаются в познавательную деятельность животного разные сенсорные и эффекторные системы. Ведь манипулирующая обезьяна почти непрерывно следит за движениями своих рук; под пристальным зрительным контролем производятся самые разнообразные действия как без разрушения целостности объекта: поворачивание в разные стороны, облизывание, поглаживание, придавливание, перекатывание и т.п., так и деструктивного порядка: разламывание, разрывание, вычленение отдельных деталей и т.д. (рис. 24).
Особую познавательную ценность представляют деструктивные действия, так как они позволяют получить сведения о внутренней структуре предметов. При манипулировании животное получает информацию одновременно по ряду сенсорных каналов, но преобладающее значение имеет у обезьян сочетание кожно-мышечной чувствительности рук со зрительными ощущениями. Кроме того, в обследовании объекта манипулирования участвуют также обоняние, вкус, тактильная чувствительность околоротовых вибрисс, иногда слух и т.д. Эти виды чувствительности сочетаются с кожно-мышечной чувствительностью эффекторов (ротового аппарата, передних конечностей) кроме обезьян и у других млекопитающих, когда те манипулируют предметами. В итоге животные получают комплексную информацию об объекте как едином целом и обладающем разнокачественньми свойствами. Именно в этом и заключается значение манипулирования как основы интеллектуального поведения.
Необходимо, однако, подчеркнуть, что первостепенное значение имеют для интеллектуального поведения зрительные восприятия и особенно зрительные обобщения, о которых уже раньше шла речь. Насколько развита способность к формированию обобщенных зрительных образов даже у крыс - показывает следующий эксперимент, в котором крысы с успехом решали очень трудную задачу: животному надо выбрать из трех предъявленных фигур (вертикальные и горизонтальные полосы) одну несходную по сравнению с двумя другими. Местонахождение и рисунок такой фигуры постоянно меняются, следовательно, это будут то вертикальные, то горизонтальные полосы, расположенные то слева, то справа, то посередине (в неправильной последовательности). Таким образом, подопытное животное могло ориентироваться лишь по одному, крайне обобщенному признаку - несхожести одного рисунка по сравнению с остальными. Мы имеем здесь, таким образом, дело со зрительным обобщением, близким к абстрагированию, свойственному мыслительным процессам.
С другим элементом интеллектуального поведения, на этот раз в двигательной сфере, мы встречались при описании опытов с «проблемными ящиками». И здесь мы имеем дело со сложными многофазовыми навыками, поскольку у высших млекопитающих, например енотов, можно сравнительно легко добиться решения задач, при которых животному приходится в определенной последовательности открывать совокупность разнообразных запирающих приспособлений. Как и при описанном опыте с подтягиванием крысой лестницы, енот может решить такую задачу лишь при соблюдении строго определенной последовательности действий. Но разница заключается в том, что в отличие от крысы еноту приходится самому находить эту последовательность, и это поднимает его деятельность, безусловно, на более высокий уровень. Правда, по некоторым данным и крысы способны на это.
Необходимо, однако, подчеркнуть, что даже высшие позвоночные решают инструментальные задачи труднее, чем локомоторные. Л. Кардош отметил в этой связи, что в психической деятельности животных преобладает познавание пространственных отношений (см. ч. I, гл. 3), постигаемых ими с помощью локомоторных действий. У обезьян же, особенно человекообразных, локомоторное познавание пространственных отношений теряет свою доминирующую роль за счет сильного развития манипуляционных действий. Однако только человек может полностью освободиться от направляющего воздействия пространственных отношений, если этого требует познание временно-причинных связей.
Решение многофазовых инструментальных задач у обезьян изучалось рядом исследователей, в частности Н. Н. Ладыгиной-Котс. В своей монографии «Приспособительные моторные навыки макака в условиях эксперимента» она подвела итог многочисленным опытам, в которых использовались весьма разнообразные сочетания запирающих механизмов. Эти эксперименты показали, что низшая обезьяна (макак-резус) способна научиться отмыкать большие серии запирающих механизмов, хотя лучше справляется с одиночными установками. Характерными явились при этом многочисленные и многообразные нащупывающие движения рук, «экспериментирование». Ввиду ее большой торопливости наиболее трудными оказались для обезьяны те приспособления, которые не отмыкались легкими быстрыми движениями. Легче всего производились такие движения, как вытягивание, отведение, притягивание, опускание и др., труднее всего - отодвигание и вращение, вывертывание. Вообще в поисках пунктов задержки и пути преодоления препятствий преобладающая роль принадлежала кинестетическим, а не зрительным восприятиям. Интересно, что по многим из этих особенностей, как показали более поздние исследования, действия низших обезьян в опытах с запирающими механизмами напоминают таковые енотов.