Страница:
«Глазки» описаны и у других жгутиковых. Например, массивный ярко-красный глазок имеет Collodictyon sparsevacuolata - своеобразный протист с 2-4 жгутиками и амебоидным обликом, способный быстро передвигаться как с помощью жгутиков, так и с помощью ложноножек. Особую сложность фоторецепция достигает у жгутикового Pauchetia (Dinoflagellata), у которого имеются уже аналоги существенных частей глаза многоклеточных животных, пигментное пятно снабжено не только светонепроницаемым экраном (аналог пигментной оболочки), но и све-топроницаемым образованием в форме сферической линзы (аналог хрусталика). Такой «глазок» позволяет не только локализовать световые лучи, но и собирать, в известной степени фокусировать их.
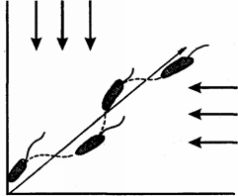
Рис. 28. Траектория передвижения эвглены под воздействием двух одинаково интенсивных источников света (по Будеру)
Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т.е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняются движения ресничек, происходит тактильное и химическое обследование объекта и т.д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.
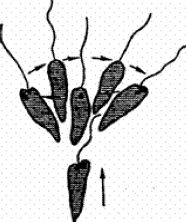 Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по
Джен-нингсу)
Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по
Джен-нингсу)
 Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф.Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю - вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф.Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю - вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.
Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).
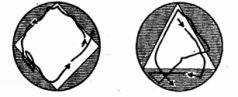
Рис. 30. Привыкание у парамеции (по Брамштедту).Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения - привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация видотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2-3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У.Грабовского).
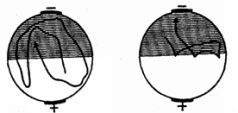
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Зесту).Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г.Зеста). Однако обратный результат, т.е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С.Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я.Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140-160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф.Б.Эпплуайт и Ф.Т.Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Общая характеристика психической активности
Как мы могли убедиться, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же мы имеем здесь дело лишь с примитивными проявлениями психической активности. О такой активности, о психике мы можем здесь говорить потому, что простейшие активно реагируют на изменения в окружающей их среде, причем реагируют на биологически непосредственно не значимые свойства компонентов среды как на сигналы о появлении жизненно важных условий среды. Другими словами, простейшим свойственна элементарная форма психического отражения - ощущение, т.е. чувствительность в собственном смысле слова. Но там, как мы уже знаем, где появляется способность к ощущению, начинается психика.
Уже говорилось и о том, что даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе: растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости. Однако элементы такого допсихического отражения встречаются и у простейших.
Большой интерес представляют в этом отношении, как и вообще для понимания условий зарождения психического отражения, реакции простейших на температуру. Здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды.
Еще в начале нашего века М.Мендельсон установил, что у инфузорий реакции на изменения температуры становятся все более дифференцированными по мере приближения к некому термическому оптимуму. Так, у Paramecium aurelia такой оптимум находится в пределах +24… +28° температуры воды. При +6… +15° инфузория реагирует на разности температуры от 0,06 до 0,08°, при +20… +24° же - на разности от 0,02 до 0,005°. Но по современным данным этот вид хорошо переносит и низкие температуры и даже замораживание до -30° и более.
Вместе с тем у простейших специфические терморецепторы, т.е. специальные образования, служащие для восприятия изменений температуры среды, не обнаружены. (Достоверно такие рецепторы неизвестны и для других беспозвоночных.) В свое время Г.Дженнингс полагал, что у инфузории (туфельки) передний конец тела особенно чувствителен в отношении температурных раздражений. Однако впоследствии О.Келер показал, что у разрезанной на две части туфельки передний и задний концы тела ведут себя в этом отношении в принципе так же, как целое животное. Отсюда напрашивается вывод, что реакция на изменения температуры является у простейших свойством всей протоплазмы. Возможно, что реакции простейших на термические раздражения еще сходны с биохимическими реакциями типа ферментативных процессов.
Очевидно, все это и соответствует тому обстоятельству, что для организма простейшего тепловая энергия еще не расчленяется на источник жизнедеятельности и сигнал о наличии такого источника. В связи с этим отсутствуют и специальные терморецепторы. Мы имеем, следовательно, здесь пример «сосуществования» у простейших допсихического и психического отражения.
Интересно, что отсутствие терморецепторов не делает реакции простейших менее выразительными, чем их реакции на другие внешние агенты. Как было показано, эти реакции проявляются в таксисах (в данном случае термотаксисах) и кинезах.
Еще явственнее «сосуществование» допсихического и психического отражения выступает у низших представителей типа. У эвглены оно обусловлено и наличием наряду с животным, растительного, аутотрофного типа питания, почему эвглена в одинаковой степени относится и к растениям и к животным.
С другой стороны, мы знаем, что качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших мы встречаем разнообразные формы передвижения в водной среде, но только на самом примитивном уровне инстинктивного поведения - кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия. За отдельными исключениями, активность простейших находится в целом как бы под отрицательным знаком, ибо эти животные попадают в сферу действия положительных раздражителей, уходя от отрицательных.
Это значит, что поисковая фаза инстинктивного поведения (кинезов) в указанном отношении еще крайне недоразвита. К тому же она явно лишена сложной, многоэтапной структуры. Возможно, что во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Ведь это содержание наполняется прежде всего активным поиском и оценкой раздражителей на первых этапах поисковой фазы.
Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. К уже упомянутым примерам положительных таксисов можно добавить, что амеба в состоянии находить пищевой объект на расстоянии до 20- 30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом: дистантно на этом уровне распознаются преимущественно отрицательные компоненты среды; биологически «нейтральные» же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные, т.е. попросту еще не существуют для животного как таковые. Таким образом, психическое отражение выполняет на самом низком уровне своего развития в основном сторожевую функцию и отличается поэтому характерной «однобокостью»: сопутствующие биологически не значимые свойства компонентов среды дистантно ощущаются животными как сигналы появления таких компонентов лишь в том случае, если эти компоненты являются для животных вредными.
Что касается, наконец, пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное научение. Последнее, как мы видели, представлено наиболее примитивной формой - привыканием, и лишь в отдельных случаях, может быть, встречаются зачатки ассоциативного научения.
Конечно, при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, во всяком случае в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов. С другой стороны, непродолжительность жизни микроорганизмов как отдельных особей (частая смена поколений) и относительное однообразие этого микромира делают излишним развитие более сложных форм накопления индивидуального опыта. В этой микросреде нет столь сложных и разнообразных условий, к которым можно приспособиться только путем научения. В таких условиях пластичность самого строения простейших, легкость образования новых морфологических структур в достаточной мере обеспечивают приспособляемость этих животных к условиям существования. Можно, очевидно, сказать, что пластичность поведения здесь еще не превзошла пластичность строения организма.
Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. Это тоже одна из причин, почему мы описали этот уровень психического развития на примере только простейших. Здесь наглядно выступает уже отмеченная общая закономерность: психологическая классификация не вполне совпадает с зоологической, так как некоторые представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, другие - уже на более высоком. Именно последнее имеет место у высших представителей типа простейших, которых в этом отношении можно было бы рассматривать как исключения. Однако, в сущности, это не так, ибо здесь проявляется и другая закономерность эволюции психики, а именно: элементы более высокого уровня психического развития всегда зарождаются в недрах предшествующего, более низкого уровня. В данном случае, например, примитивные формы ассоциативного научения, вообще характерные для более высокого уровня элементарной сенсорной психики, встречаются в зачатке уже у некоторых видов, относящихся к типу, который в целом стоит на низшем уровне элементарной сенсорной психики, где типичной формой индивидуально-изменчивого поведения является привыкание.
Высший уровень развития элементарной сенсорной психики
Рис. 28. Траектория передвижения эвглены под воздействием двух одинаково интенсивных источников света (по Будеру)
Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т.е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняются движения ресничек, происходит тактильное и химическое обследование объекта и т.д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.
Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по
Джен-нингсу)
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф.Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю - вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).
Рис. 30. Привыкание у парамеции (по Брамштедту).Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения - привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация видотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2-3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У.Грабовского).
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Зесту).Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г.Зеста). Однако обратный результат, т.е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С.Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я.Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140-160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф.Б.Эпплуайт и Ф.Т.Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Общая характеристика психической активности
Как мы могли убедиться, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же мы имеем здесь дело лишь с примитивными проявлениями психической активности. О такой активности, о психике мы можем здесь говорить потому, что простейшие активно реагируют на изменения в окружающей их среде, причем реагируют на биологически непосредственно не значимые свойства компонентов среды как на сигналы о появлении жизненно важных условий среды. Другими словами, простейшим свойственна элементарная форма психического отражения - ощущение, т.е. чувствительность в собственном смысле слова. Но там, как мы уже знаем, где появляется способность к ощущению, начинается психика.
Уже говорилось и о том, что даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе: растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости. Однако элементы такого допсихического отражения встречаются и у простейших.
Большой интерес представляют в этом отношении, как и вообще для понимания условий зарождения психического отражения, реакции простейших на температуру. Здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды.
Еще в начале нашего века М.Мендельсон установил, что у инфузорий реакции на изменения температуры становятся все более дифференцированными по мере приближения к некому термическому оптимуму. Так, у Paramecium aurelia такой оптимум находится в пределах +24… +28° температуры воды. При +6… +15° инфузория реагирует на разности температуры от 0,06 до 0,08°, при +20… +24° же - на разности от 0,02 до 0,005°. Но по современным данным этот вид хорошо переносит и низкие температуры и даже замораживание до -30° и более.
Вместе с тем у простейших специфические терморецепторы, т.е. специальные образования, служащие для восприятия изменений температуры среды, не обнаружены. (Достоверно такие рецепторы неизвестны и для других беспозвоночных.) В свое время Г.Дженнингс полагал, что у инфузории (туфельки) передний конец тела особенно чувствителен в отношении температурных раздражений. Однако впоследствии О.Келер показал, что у разрезанной на две части туфельки передний и задний концы тела ведут себя в этом отношении в принципе так же, как целое животное. Отсюда напрашивается вывод, что реакция на изменения температуры является у простейших свойством всей протоплазмы. Возможно, что реакции простейших на термические раздражения еще сходны с биохимическими реакциями типа ферментативных процессов.
Очевидно, все это и соответствует тому обстоятельству, что для организма простейшего тепловая энергия еще не расчленяется на источник жизнедеятельности и сигнал о наличии такого источника. В связи с этим отсутствуют и специальные терморецепторы. Мы имеем, следовательно, здесь пример «сосуществования» у простейших допсихического и психического отражения.
Интересно, что отсутствие терморецепторов не делает реакции простейших менее выразительными, чем их реакции на другие внешние агенты. Как было показано, эти реакции проявляются в таксисах (в данном случае термотаксисах) и кинезах.
Еще явственнее «сосуществование» допсихического и психического отражения выступает у низших представителей типа. У эвглены оно обусловлено и наличием наряду с животным, растительного, аутотрофного типа питания, почему эвглена в одинаковой степени относится и к растениям и к животным.
С другой стороны, мы знаем, что качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших мы встречаем разнообразные формы передвижения в водной среде, но только на самом примитивном уровне инстинктивного поведения - кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия. За отдельными исключениями, активность простейших находится в целом как бы под отрицательным знаком, ибо эти животные попадают в сферу действия положительных раздражителей, уходя от отрицательных.
Это значит, что поисковая фаза инстинктивного поведения (кинезов) в указанном отношении еще крайне недоразвита. К тому же она явно лишена сложной, многоэтапной структуры. Возможно, что во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Ведь это содержание наполняется прежде всего активным поиском и оценкой раздражителей на первых этапах поисковой фазы.
Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. К уже упомянутым примерам положительных таксисов можно добавить, что амеба в состоянии находить пищевой объект на расстоянии до 20- 30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом: дистантно на этом уровне распознаются преимущественно отрицательные компоненты среды; биологически «нейтральные» же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные, т.е. попросту еще не существуют для животного как таковые. Таким образом, психическое отражение выполняет на самом низком уровне своего развития в основном сторожевую функцию и отличается поэтому характерной «однобокостью»: сопутствующие биологически не значимые свойства компонентов среды дистантно ощущаются животными как сигналы появления таких компонентов лишь в том случае, если эти компоненты являются для животных вредными.
Что касается, наконец, пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное научение. Последнее, как мы видели, представлено наиболее примитивной формой - привыканием, и лишь в отдельных случаях, может быть, встречаются зачатки ассоциативного научения.
Конечно, при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, во всяком случае в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов. С другой стороны, непродолжительность жизни микроорганизмов как отдельных особей (частая смена поколений) и относительное однообразие этого микромира делают излишним развитие более сложных форм накопления индивидуального опыта. В этой микросреде нет столь сложных и разнообразных условий, к которым можно приспособиться только путем научения. В таких условиях пластичность самого строения простейших, легкость образования новых морфологических структур в достаточной мере обеспечивают приспособляемость этих животных к условиям существования. Можно, очевидно, сказать, что пластичность поведения здесь еще не превзошла пластичность строения организма.
Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. Это тоже одна из причин, почему мы описали этот уровень психического развития на примере только простейших. Здесь наглядно выступает уже отмеченная общая закономерность: психологическая классификация не вполне совпадает с зоологической, так как некоторые представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, другие - уже на более высоком. Именно последнее имеет место у высших представителей типа простейших, которых в этом отношении можно было бы рассматривать как исключения. Однако, в сущности, это не так, ибо здесь проявляется и другая закономерность эволюции психики, а именно: элементы более высокого уровня психического развития всегда зарождаются в недрах предшествующего, более низкого уровня. В данном случае, например, примитивные формы ассоциативного научения, вообще характерные для более высокого уровня элементарной сенсорной психики, встречаются в зачатке уже у некоторых видов, относящихся к типу, который в целом стоит на низшем уровне элементарной сенсорной психики, где типичной формой индивидуально-изменчивого поведения является привыкание.
Высший уровень развития элементарной сенсорной психики
Высшего уровня элементарной сенсорной психики достигло большое число многоклеточных беспозвоночных. Однако, как отмечалось, часть низших многоклеточных беспозвоночных находится в основном на том же уровне психического развития, что и многие простейшие. Это относится прежде всего к большинству кишечнополостных и к низшим червям, а тем более к губкам, которые во многом еще напоминают колониальные формы одноклеточных (жгутиковых). Неподвижный, сидячий образ жизни взрослых губок привел даже к редукции их внешней активности, поведения (при полном отсутствии нервной системы и органов чувств). Но даже у самых примитивных представителей многоклеточных животных создались принципиально новые условия поведения в результате появления качественно новых структурных категорий - тканей, органов, систем органов. Это и обусловило возникновение специальной системы координации деятельности этих многоклеточных образований и усложнившегося взаимодействия организма со средой - нервной системы.
К низшим многоклеточным беспозвоночным относятся помимо уже упомянутых еще иглокожие, высшие (кольчатые) черви, отчасти моллюски и др.
Мы рассмотрим в дальнейшем в качестве примера кольчатых червей, у которых в полной мере выражены признаки поведения, характерные для высшего уровня элементарной сенсорной психики. К кольчатым червям относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви (наиболее известный представитель - дождевой червь) и пиявки. Характерным признаком строения кольчецов является внешняя и внутренняя метамерия: тело состоит из нескольких большей частью идентичных сегментов, каждый из которых содержит «комплект» внутренних органов, в частности пару симметрично расположенных ганглиев с нервными коммисурами. В результате нервная система кольчатых червей имеет вид «нервной лестницы».
Нервная система
Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы - важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Важным моментом здесь является уже резкое ускорение проводимости возбуждения в нервной ткани: упротоплазме скорость проведения возбуждения не превышает 1-2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших (бескишечных) плоских червей и примитивных моллюсков (хитон) еще только сетью, располагающейся вблизи поверхности тела, но выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова (процесс цефализации), а вместе с ней и головной мозг - скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение «нервной лестницы», при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммисурами («окологлоточное кольцо») с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому говорят и о ганглионарной нервной системе, или о «ганглионарной лестнице». У некоторых представителей данной группы животных (например, пиявок) нервные стволы сближаются настолько, что получается «нервная цепочка».
К низшим многоклеточным беспозвоночным относятся помимо уже упомянутых еще иглокожие, высшие (кольчатые) черви, отчасти моллюски и др.
Мы рассмотрим в дальнейшем в качестве примера кольчатых червей, у которых в полной мере выражены признаки поведения, характерные для высшего уровня элементарной сенсорной психики. К кольчатым червям относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви (наиболее известный представитель - дождевой червь) и пиявки. Характерным признаком строения кольчецов является внешняя и внутренняя метамерия: тело состоит из нескольких большей частью идентичных сегментов, каждый из которых содержит «комплект» внутренних органов, в частности пару симметрично расположенных ганглиев с нервными коммисурами. В результате нервная система кольчатых червей имеет вид «нервной лестницы».
Нервная система
Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы - важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Важным моментом здесь является уже резкое ускорение проводимости возбуждения в нервной ткани: упротоплазме скорость проведения возбуждения не превышает 1-2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших (бескишечных) плоских червей и примитивных моллюсков (хитон) еще только сетью, располагающейся вблизи поверхности тела, но выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова (процесс цефализации), а вместе с ней и головной мозг - скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение «нервной лестницы», при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммисурами («окологлоточное кольцо») с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому говорят и о ганглионарной нервной системе, или о «ганглионарной лестнице». У некоторых представителей данной группы животных (например, пиявок) нервные стволы сближаются настолько, что получается «нервная цепочка».