Страница:
От ганглиев отходят мощные проводящие волокна, которые и составляют нервные стволы. В гигантских волокнах нервные импульсы проводятся значительно быстрее благодаря их большому диаметру и малому числу синаптических связей (мест соприкосновения аксонов одних нервных клеток с дендритами и клеточными телами других клеток). Что же касается головных ганглиев, т.е. мозга, то они больше развиты у более подвижных животных, обладающих и наиболее развитыми рецепторными системами.
Зарождение и эволюция нервной системы обусловлены необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
Огромное значение имел в этом отношении и процесс цефализапии, т.е. обособления головного конца организма и сопряженного с ним появления головного мозга. Только при наличии головного мозга возможно подлинно централизованное «кодирование» поступающих с периферии сигналов и формирование целостных «программ» врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животного.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Это опять показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. Одновременно этот пример еще раз демонстрирует, с какой осторожностью надо подходить к оценке «высших» и «низших» признаков при сравнении организмов, занимающих различное филогенетическое положение, в частности при сопоставлении простейших и многоклеточных беспозвоночных.
Движения
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Хищные виды отправляются из этих убежищ регулярно на «охоту». Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупалец, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободноживущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малощетинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается детритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки подстерегают свои жертвы на суше (млекопитающих и людей), в траве или на ветках деревьев и кустарников (во влажных тропических лесах). Эти пиявки могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает сперва туловище, затем присасывается к субстрату головной присоской, притягивает к ней задний конец туловища (с одновременным сокращением последнего), присасывается задней присоской и т.д.
Итак, двигательная активность кольчатых червей как при локомоции, так и при добывании пищи отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, представленной прежде всего так называемым кожно-мышечным мешком. Он состоит из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают локомоцию: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается (и сужается) передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают «волны» сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: кольчецы (помимо пиявок) носят на каждом сегменте по паре выростов, служащих органами передвижения (кроме головного конца, где они служат ротовыми органами) и получивших название параподий. Параподии снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподий имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке - челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многошетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподии служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и представлены в большем количестве.
Органы чувств и сенсорные способности
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т.д. Очевидно, первичные органы чувств вообще были плюромодальными, т.е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромодальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания - наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или веретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функции слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюромодальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания - так называемые «сенсиллы» - превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных B-А.Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных - медузы - обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело - студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
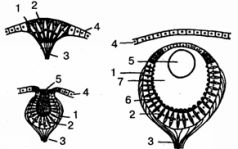
Рис. 32. Развитие глаза беспозвоночных (на примере гидромедуз). Слева вверху - глазное пятно; слева внизу - глазная ямка; справа - глазной пузырь. 1 -зрительная клетка; 2 -пигментная клетка; 3 - зрительный нерв; 4 - эпителий; 5 - хрусталик; 6 - светочувствительные палочки; 7-стекловидное тело (по Бючли)
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3%. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1%).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию светоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом - втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела - в наибольшей мере червь реагирует при освещении всей его поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) - на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие рецепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к пларающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Таксисы
Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и требования к локализации биологически значимых компонентов среды по их биологически незначимым признакам. Возникает необходимость более сложного таксисного поведения, позволяющего животному достаточно четко и дифференцированно ощущать и реагировать. Именно этим и отличаются животные, находящиеся на рассматриваемом здесь уровне развития психики: благодаря симметричному расположению органов чувств у них обнаруживаются наряду с кинезами и элементарными таксисами и некоторые высшие формы таксисного поведения.
Немецкий ученый А.Кюн выделил следующие категории высших таксисов, которые, правда, вполне развиты лишь у высших животных: тропотаксисы -движение ориентируется по равнодействующей, образуемой в результате выравнивания интенсивности возбуждения в симметрично расположенных рецепторах; телотаксисы -выбор и фиксация одного источника раздражения и направление движения к этому источнику («цели»); менотаксисы- при несимметричном раздражении в симметрично расположенных рецепторах движение производится под углом к источнику раздражения. Менотаксисы играют ведущую роль в сохранении животным константного положения в пространстве.
Кроме того, у высших животных с развитой памятью встречается еще одна форма таксисов - мнемотаксисы,при которых основную роль играет индивидуальное запоминание ориентиров, что особенно важно для территориального поведения, о чем еще пойдет речь.
В поведении кольчецов тропо- и телотаксисы чаще всего проявляются совместно: если животное подвергнуть одновременному воздействию двух источников энергии (например, света) одинаковой силы, то при положительном таксисе оно будет первоначально двигаться по направлению равнодействующей, т.е. линии, ведущей к середине расстояния между источниками (тропотаксис), но затем, как правило, еще не дойдя до этой точки, оно повернет и направится к одному из источников энергии (телотаксис).
Наиболее четко тропо- и телотаксисы проявляются у активно двигающихся кольчатых червей, прежде всего у хищников. Последние не только обнаруживают присутствие жертвы по исходящим от нее физическим и химическим стимулам, но и направляются к ней, ориентируясь на основе таких таксисов. Наземные пиявки, например, способны точно локализовать местонахождение жертвы и с удивительной быстротой направляются к ней, ориентируясь по сотрясению субстрата, а затем, на более близком расстоянии, и по исходящему от жертвы (крупных млекопитающих) теплу и запаху. В результате хищник обнаруживает ее по соответствующим градиентам с помощью сочетания положительных вибрационных, термических и химических телотаксисов.
При менотаксисах животное, как говорилось, двигается под углом к линии воздействия, исходящей от источника энергии. Типичный пример - ориентация мигрирующих животных по солнечному (или другому астрономическому) «компасу», что, конечно, особенно характерно для перелетных птиц. У низших многоклеточных беспозвоночных менотаксисы обнаружены у некоторых улиток. Если этих животных поместить на освещаемый сбоку и медленно вращающийся диск, то они, двигаясь против направления вращения, придерживаются определенного угла по отношению к источнику света. В этих случаях, следовательно, можно уже говорить об ориентации животного по световому компасу, которая играет исключительную роль в жизни высших животных.
Зачатки высших форм поведения
Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов психической деятельности, присущей более высокоорганизованным животным. В отличие от основной массы низших беспозвоночных у полихет наблюдаются некоторые существенные усложнения видотипичного поведения, отчасти уже выходящие за рамки типичной элементарной сенсорной психики.
Так, в некоторых случаях у этих морских червей встречаются действия, которые уже можно назвать конструктивными, поскольку животные активно создают сооружения из отдельных посторонних частиц, скрепляя их в единое целое. Речь идет о постройке уже упомянутых «домиков» - трубок из отдельных частичек, которые собираются червями на морском дне и укрепляются с помощью специальных «рабочих» органов - преобразованных передних параподий.
Сам процесс сооружения трубки является сложной деятельностью, состоящей из нескольких фаз, адекватно видоизменяющихся в зависимости от таких внешних факторов, как характер грунта и течения, рельеф дна, количество и состав опускающихся на дно частиц, скорость их оседания и т.д. Строительным материалом служат разного рода мелкий обломочный материал (например, кусочки раковин), песчинки, частицы растений и т.п., которые скрепляются специальными выделениями особого «цементирующего органа». Пригодные для употребления частицы выбираются животным, причем в зависимости от его возраста; молодые особи используют лишь мелкие гранулы, более старые - более крупные. Все частицы собираются и прикрепляются друг к другу уже упомянутыми специализированными щупальцами. Полихета Aulophorus carteri, например, строит трубки из спор водных растений, которые она собирает и прикрепляет друг к другу вокруг себя подобно тому, как строится труба из кирпичей (рис.33).
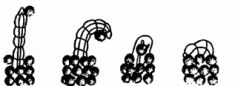
Рис. 33. Постройка многощетинковым червем трубки из спор водных растений (по Грассэ)
Не меньший интерес представляют впервые намечающиеся у многощетинковых червей проявления брачного поведения и агрессивности, а вместе с ними и элементов общения. Конечно, подлинные агрессивность и брачное поведение характеризуются ритуализацией, которая появляется только у головоногих моллюсков и членистоногих, т.е. на низшем уровне перцептивной психики. Все же у представителей рода Nereis удалось наблюдать борьбу между двумя червями. У Nereis pelagica такая борьба может начаться (но не обязательно), если одна особь пытается проникнуть в «домик» другой особи. Иногда вторгшаяся особь кусает владельца «домика» в задний конец тела, и тогда «хозяин» покидает трубку или же, перевернувшись, вступает в борьбу с пришельцем. В других случаях, когда животные встречаются в трубке на полпути, головой к голове, борьба вспыхивает немедленно, и черви борются, расположившись брюшными сторонами и головами друг к другу. Однако при всех обстоятельствах животные не наносят друг другу повреждения. В более ожесточенной, «непримиримой» форме борьба происходит у Nereis caudata, причем не только из-за обладания «домиком». «Драки» описаны английским исследователем поведения морских кольчатых червей С.М.Эвансом и его сотрудниками также у полихеты Harmothoe imbricata. У этого вида борьба может возникнуть при случайной встрече двух особей, особенно же агрессивность проявляется в сфере воспроизведения, при образовании пар: самец, уцепившийся за самку, становится чрезвычайно агрессивным по отношению к другим самцам (но не самкам). Никогда, однако, борьба не сопровождается у полихет подачей каких-либо сигналов или другими проявлениями ритуализации поведения.
Зарождение и эволюция нервной системы обусловлены необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
Огромное значение имел в этом отношении и процесс цефализапии, т.е. обособления головного конца организма и сопряженного с ним появления головного мозга. Только при наличии головного мозга возможно подлинно централизованное «кодирование» поступающих с периферии сигналов и формирование целостных «программ» врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животного.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Это опять показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. Одновременно этот пример еще раз демонстрирует, с какой осторожностью надо подходить к оценке «высших» и «низших» признаков при сравнении организмов, занимающих различное филогенетическое положение, в частности при сопоставлении простейших и многоклеточных беспозвоночных.
Движения
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Хищные виды отправляются из этих убежищ регулярно на «охоту». Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупалец, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободноживущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малощетинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается детритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки подстерегают свои жертвы на суше (млекопитающих и людей), в траве или на ветках деревьев и кустарников (во влажных тропических лесах). Эти пиявки могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает сперва туловище, затем присасывается к субстрату головной присоской, притягивает к ней задний конец туловища (с одновременным сокращением последнего), присасывается задней присоской и т.д.
Итак, двигательная активность кольчатых червей как при локомоции, так и при добывании пищи отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, представленной прежде всего так называемым кожно-мышечным мешком. Он состоит из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают локомоцию: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается (и сужается) передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают «волны» сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: кольчецы (помимо пиявок) носят на каждом сегменте по паре выростов, служащих органами передвижения (кроме головного конца, где они служат ротовыми органами) и получивших название параподий. Параподии снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподий имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке - челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многошетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподии служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и представлены в большем количестве.
Органы чувств и сенсорные способности
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т.д. Очевидно, первичные органы чувств вообще были плюромодальными, т.е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромодальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания - наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или веретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функции слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюромодальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания - так называемые «сенсиллы» - превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных B-А.Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных - медузы - обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело - студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
Рис. 32. Развитие глаза беспозвоночных (на примере гидромедуз). Слева вверху - глазное пятно; слева внизу - глазная ямка; справа - глазной пузырь. 1 -зрительная клетка; 2 -пигментная клетка; 3 - зрительный нерв; 4 - эпителий; 5 - хрусталик; 6 - светочувствительные палочки; 7-стекловидное тело (по Бючли)
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3%. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1%).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию светоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом - втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела - в наибольшей мере червь реагирует при освещении всей его поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) - на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие рецепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к пларающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Таксисы
Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и требования к локализации биологически значимых компонентов среды по их биологически незначимым признакам. Возникает необходимость более сложного таксисного поведения, позволяющего животному достаточно четко и дифференцированно ощущать и реагировать. Именно этим и отличаются животные, находящиеся на рассматриваемом здесь уровне развития психики: благодаря симметричному расположению органов чувств у них обнаруживаются наряду с кинезами и элементарными таксисами и некоторые высшие формы таксисного поведения.
Немецкий ученый А.Кюн выделил следующие категории высших таксисов, которые, правда, вполне развиты лишь у высших животных: тропотаксисы -движение ориентируется по равнодействующей, образуемой в результате выравнивания интенсивности возбуждения в симметрично расположенных рецепторах; телотаксисы -выбор и фиксация одного источника раздражения и направление движения к этому источнику («цели»); менотаксисы- при несимметричном раздражении в симметрично расположенных рецепторах движение производится под углом к источнику раздражения. Менотаксисы играют ведущую роль в сохранении животным константного положения в пространстве.
Кроме того, у высших животных с развитой памятью встречается еще одна форма таксисов - мнемотаксисы,при которых основную роль играет индивидуальное запоминание ориентиров, что особенно важно для территориального поведения, о чем еще пойдет речь.
В поведении кольчецов тропо- и телотаксисы чаще всего проявляются совместно: если животное подвергнуть одновременному воздействию двух источников энергии (например, света) одинаковой силы, то при положительном таксисе оно будет первоначально двигаться по направлению равнодействующей, т.е. линии, ведущей к середине расстояния между источниками (тропотаксис), но затем, как правило, еще не дойдя до этой точки, оно повернет и направится к одному из источников энергии (телотаксис).
Наиболее четко тропо- и телотаксисы проявляются у активно двигающихся кольчатых червей, прежде всего у хищников. Последние не только обнаруживают присутствие жертвы по исходящим от нее физическим и химическим стимулам, но и направляются к ней, ориентируясь на основе таких таксисов. Наземные пиявки, например, способны точно локализовать местонахождение жертвы и с удивительной быстротой направляются к ней, ориентируясь по сотрясению субстрата, а затем, на более близком расстоянии, и по исходящему от жертвы (крупных млекопитающих) теплу и запаху. В результате хищник обнаруживает ее по соответствующим градиентам с помощью сочетания положительных вибрационных, термических и химических телотаксисов.
При менотаксисах животное, как говорилось, двигается под углом к линии воздействия, исходящей от источника энергии. Типичный пример - ориентация мигрирующих животных по солнечному (или другому астрономическому) «компасу», что, конечно, особенно характерно для перелетных птиц. У низших многоклеточных беспозвоночных менотаксисы обнаружены у некоторых улиток. Если этих животных поместить на освещаемый сбоку и медленно вращающийся диск, то они, двигаясь против направления вращения, придерживаются определенного угла по отношению к источнику света. В этих случаях, следовательно, можно уже говорить об ориентации животного по световому компасу, которая играет исключительную роль в жизни высших животных.
Зачатки высших форм поведения
Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов психической деятельности, присущей более высокоорганизованным животным. В отличие от основной массы низших беспозвоночных у полихет наблюдаются некоторые существенные усложнения видотипичного поведения, отчасти уже выходящие за рамки типичной элементарной сенсорной психики.
Так, в некоторых случаях у этих морских червей встречаются действия, которые уже можно назвать конструктивными, поскольку животные активно создают сооружения из отдельных посторонних частиц, скрепляя их в единое целое. Речь идет о постройке уже упомянутых «домиков» - трубок из отдельных частичек, которые собираются червями на морском дне и укрепляются с помощью специальных «рабочих» органов - преобразованных передних параподий.
Сам процесс сооружения трубки является сложной деятельностью, состоящей из нескольких фаз, адекватно видоизменяющихся в зависимости от таких внешних факторов, как характер грунта и течения, рельеф дна, количество и состав опускающихся на дно частиц, скорость их оседания и т.д. Строительным материалом служат разного рода мелкий обломочный материал (например, кусочки раковин), песчинки, частицы растений и т.п., которые скрепляются специальными выделениями особого «цементирующего органа». Пригодные для употребления частицы выбираются животным, причем в зависимости от его возраста; молодые особи используют лишь мелкие гранулы, более старые - более крупные. Все частицы собираются и прикрепляются друг к другу уже упомянутыми специализированными щупальцами. Полихета Aulophorus carteri, например, строит трубки из спор водных растений, которые она собирает и прикрепляет друг к другу вокруг себя подобно тому, как строится труба из кирпичей (рис.33).
Рис. 33. Постройка многощетинковым червем трубки из спор водных растений (по Грассэ)
Не меньший интерес представляют впервые намечающиеся у многощетинковых червей проявления брачного поведения и агрессивности, а вместе с ними и элементов общения. Конечно, подлинные агрессивность и брачное поведение характеризуются ритуализацией, которая появляется только у головоногих моллюсков и членистоногих, т.е. на низшем уровне перцептивной психики. Все же у представителей рода Nereis удалось наблюдать борьбу между двумя червями. У Nereis pelagica такая борьба может начаться (но не обязательно), если одна особь пытается проникнуть в «домик» другой особи. Иногда вторгшаяся особь кусает владельца «домика» в задний конец тела, и тогда «хозяин» покидает трубку или же, перевернувшись, вступает в борьбу с пришельцем. В других случаях, когда животные встречаются в трубке на полпути, головой к голове, борьба вспыхивает немедленно, и черви борются, расположившись брюшными сторонами и головами друг к другу. Однако при всех обстоятельствах животные не наносят друг другу повреждения. В более ожесточенной, «непримиримой» форме борьба происходит у Nereis caudata, причем не только из-за обладания «домиком». «Драки» описаны английским исследователем поведения морских кольчатых червей С.М.Эвансом и его сотрудниками также у полихеты Harmothoe imbricata. У этого вида борьба может возникнуть при случайной встрече двух особей, особенно же агрессивность проявляется в сфере воспроизведения, при образовании пар: самец, уцепившийся за самку, становится чрезвычайно агрессивным по отношению к другим самцам (но не самкам). Никогда, однако, борьба не сопровождается у полихет подачей каких-либо сигналов или другими проявлениями ритуализации поведения.