Страница:
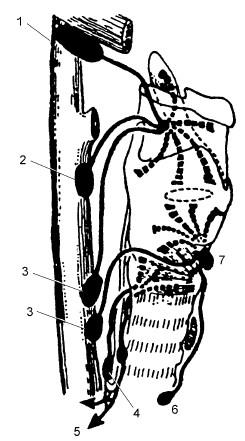
Рис. 18.2. Лимфатическая система гортани; вид справа:
1 , 2, 3 – верхние, средние и нижние яремные лимфатические узлы; 4 – лимфатические узлы, сопровождающие возвратный нерв; 5 – направления к лимфатическим узлам средостения; 6 – претрахеальные лимфатические узлы; 7 – предгортанные лимфатические узлы
Возвратный гортанный нерв (n. laryngeus recurrens) (рис. 18.3, 15), как и верхний гортанный нерв, содержит чувствительные (кроме вкусовых), двигательные и парасимпатические волокна. Правый возвратный нерв отходит от блуждающего нерва на уровне его пересечения с подключичной артерией, левый возвратный нерв отходит от блуждающего нерва на уровне его пересечения с дугой аорты латеральнее артериальной связки (lig. arteriosum). Оба возвратных нерва, огибая сзади правый – подключичную артерию, левый – дугу аорты, поднимаются спереди от них кверху, первый – по боковой поверхности трахеи, второй – в борозде между трахеей и выступающим из-под нее пищеводом. Далее оба нерва, каждый на своей стороне, пересекаются у нижнего края щитовидной железы с нижней щитовидной артерией (a. thyreoidea inferior) и подходят к гортани уже как нижние гортанные нервы. Возвратные нервы отдают следующие ветви: нижние шейные сердечные нервы (16); трахеальные ветви (17), иннервирующие слизистую оболочку, железы и гладкие мышцы трахеи; пищеводные ветви (18), иннервирующие слизистую оболочку, железы и поперечнополосатую мускулатуру верхнего отдела пищевода. Нижний гортанный нерв (n. laryngeus inferior), завершающий собою возвратный нерв, иннервирует все мышцы гортани, кроме перстнещитовидной, слизистую оболочку гортани ниже голосовой щели, а также щитовидную и паращитовидную железы и лимфоузлы шеи. Нижний гортанный нерв имеет многочисленные связи с симпатическими узлами, сосудистыми нервными сплетениями и с верхним гортанным нервом.
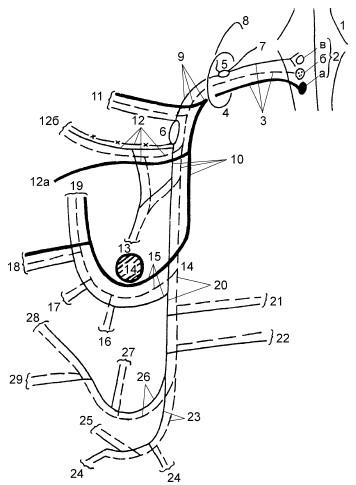
Рис. 18.3. Схема блуждающего нерва. Объяснения в тексте
Точно не известно, где у человека располагаются кортикальные зоны произвольных движений мышц гортани. Предполагают, что непарный центр речедвигательного аппарата (двигательный центр речи Брока[1]) находится в задней части нижней лобной извилины, у правшей – в левом полушарии, у левшей – в правом полушарии (рис. 18.4, 3).
Этот центр имеет тесные связи с ядром устной речи (5) звукового анализатора (центр Вернике[2]), расположенным в задней части верхней височной извилины (gyr. temporalis superior), в глубине латеральной борозды. Депривация центра Вернике, возникающая в результате ранней глухоты до приобретения ребенком речедвигательных навыков, обусловливает возникновение немоты, т. е. афункциональность двигательного центра речи Брока.
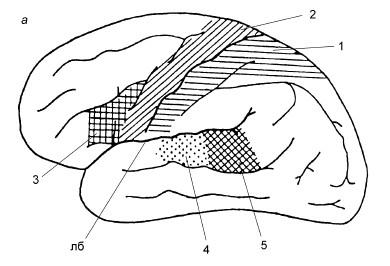
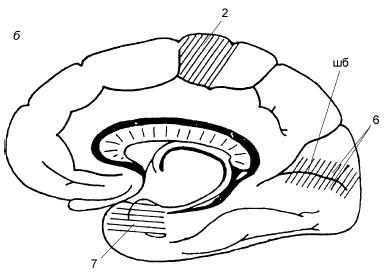
Рис. 18.4. Схема расположения корковых концов анализаторов:
а – верхнелатеральная поверхность левого полушария; б – медиальная поверхность правого полушария; 1 – ядро кожного анализатора (тактильная, болевая, температурная чувствительность); 2 – ядро двигательного анализатора; расположено в предцентральной извилине и в верхней теменной дольке; 3 – речедвигательный анализатор; расположен в задней части нижней лобной извилины (двигательный центр речи Брока, односторонний – у правшей в левом полушарии, у левшей – в правом); 4 – ядро звукового анализатора; расположено в задней части верхней височной извилины на поверхности, обращенной к островку – поперечные височные извилины; 5 – ядро звукового анализатора устной речи; расположено в задней части верхней височной извилины, в глубине латеральной борозды (лб) – центр речи Вернике; 6 – ядро зрительного анализатора; расположено по краям шпорной борозды (шб); 7 – ядро обонятельного и вкусового анализаторов; расположено в крючке
Двигательная функция гортани обеспечена двусторонней иннервацией. Корковые центры осуществляют только функцию приведения голосовых складок, поэтому в опытах на животных одностороннее разрушение двигательного центра гортани не приводит к параличу ее аддукторов. У людей даже при полной гемиплегии, возникающей при кровоизлиянии во внутреннюю капсулу, не возникает паралич внутренних мышц гортани. Однако при одностороннем ядерном поражении возникает их односторонний периферический паралич.
Корково-ядерный пирамидный путь (рис. 18.5, 4), обеспечивающий произвольные движения внутренних мышц гортани, начинается во внутренних больших пирамидных нейронах коры предцентральной извилины; он огибает полуовальный центр (2) и в составе внутренней капсулы (3) вступает в мост и далее в продолговатый мозг. Здесь отдает волокна в двигательные ядра своей и противоположной стороны, осуществляя перекрест пирамидного пути. Кроме пирамидных волокон двигательную функцию гортани обеспечивают экстрапирамидные волокна, которые берут начало в подкорковых центрах и осуществляют рефлекторную (непроизвольную) двигательную функцию гортани. К этим центрам относятся базальные ядра полушарий, зрительные бугры, красные ядра, черное вещество, ядра оливы. Переключение рефлекторных дуг гортани с восходящих (сенсорных) на нисходящие (моторные) пути происходит в указанных центрах. К экстрапирамидным путям относятся: красноядерно-спинномозговой, покрышечно-спинномозговой, сетевидно-спинномозговой и ряд других. Большинство из них направляют свои волокна к ядрам двигательных черепных нервов, в том числе и гортанных. Они обеспечивают автоматическое включение соответствующих мышц при различной сенсорной стимуляции чувствительных нервов и поддерживают тонус поперечно-полосатой мускулатуры, являющийся важным функциональным и трофическим фактором в обеспечении ее функциональной готовности. Тонус внутренних мышц гортани обеспечивает их непрерывную деятельность, оперативное включение для выполнения защитной функции, а также реципрокное расслабление и напряжение сжимателей и расширителей гортани, синхронизирующееся с фазами вдоха и выдоха (расширение при вдохе и сужение при выдохе). Пирамидная и экстрапирамидная системы в отношении функций глотки и гортани находятся в состоянии постоянного взаимодействия, синхронизирующегося со сложной афферентацией, поступающей из рецепторов этих анатомических образований.
На рис. 18.6 приведена схема гортанных нервов и их многочисленных разветвлений. Чувствительная иннервация гортани полностью обеспечивается верхним гортанным нервом. Что касается двигательной функции возвратного гортанного нерва, то приблизительно 70–75 % его волокон иннервируют сжиматели гортани и лишь 25–30 % – единственную пару расширителей – перстнечерпаловидную мышцу (m. crico-arytenoidei), поэтому при поражении волокон возвратного нерва могут возникать следующие варианты положения голосовой складки: при тотальном поражении возникает так называемое трупное положение, среднее между приведением и отведением; при поражении ветви, иннервирующей расширитель гортани, голосовая складка занимает срединное положение за счет действия мощной системы констрикторов.
Единственная пара мышц, не иннервируемая возвратным нервом, – это перстнещитовидные мышцы. Все остальные внутренние гортанные мышцы иннервируются волокнами возвратного нерва. Эти волокна, достигнув гортани, распространяются под слизистой оболочкой, покрывающей ее заднюю поверхность, и распределяются среди всех мышц гортани, исключая перстнещитовидную, иннервируемую наружной ветвью верхнего гортанного нерва. Возвратный нерв отдает единственную чувствительную ветвь, которая анастомозирует на задней поверхности гортани с чувствительной ветвью внутренней ветви верхнего гортанного нерва, образуя петлю Галена (5), располагающуюся под слизистой оболочкой на мышцах, покрывающих заднюю поверхность перстневидного хряща (5).
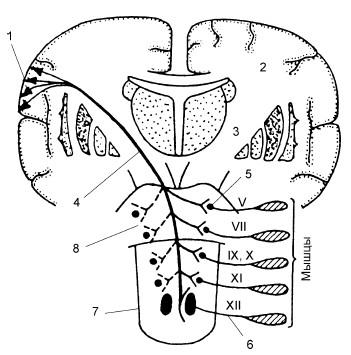
Рис. 18.5. Корково-ядерный пирамидный путь:
1 – пирамидные клетки (тела первых нейронов); 2 – полуовальный центр; 3 – внутренняя капсула; 4 – корково-ядерный путь; 5 – тела вторых нейронов; 6 – аксон второго нейрона; 7 – продолговатый мозг; 8 – мост; V, VII, IX, X, XI, XII – пары черепных нервов
Трофическое влияние на гортань оказывают симпатические волокна, исходящие из шейного симпатического ствола. Эти же волокна оказывают сосудорасширяющее действие и, по-видимому, участвуют в поддержании тонуса голосовых складок (голосовой мышцы).
Значительная протяженность возвратных нервов, их тесное соседство с различными органами шеи (щитовидная железа, трахея, дуга аорты, лимфоузлы, пищевод и др.) обусловливают их частое поражение при различных патологических состояниях указанных органов и анатомических образований.
Физиология гортани
Гортань обеспечивает ряд функций (фонаторную, защитную, включающую запирательную, кашлевую, отхаркивающую, опосредованную функцию фиксации плечевого пояса при физическом усилии и оптимизацию дыхательной функции легких), а также дыхательную и выделившуюся в процессе эволюции специфическую для человека – речеобразовательную, основанную на фонаторной и дыхательной функциях.
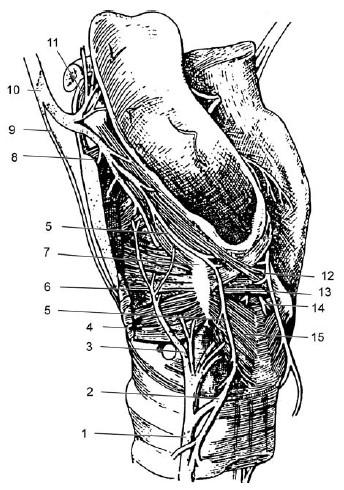
Рис. 18.6. Нервы гортани, вид слева:
1 – возвратный нерв; 2 – нижний сегмент петли Галена[3]; 3, 6 – волокна к щиточерпаловидной мышце; 4 – латеральная перстнечерпаловидная мышца; 5 – петля Галена; 7 – черпаловидный хрящ; 8 – продолжение верхнего гортанного нерва; 9 – наружная ветвь верхнего гортанного нерва; 10 – ствол верхнего гортанного нерва; 11 – нервные волокна к надгортаннику; 12, 13 – косые и поперечные волокна черпало-черпаловидной мышцы; 14 – нервные волокна к черпало-черпаловидной мышце; 15 – задняя перстнечерпаловидная мышца
Дыхательная функция гортани заключается не в пассивном пропускании воздуха в легкие и обратно, а в ряде активных рефлекторных актов, совершаемых в гортани и оптимизирующих легочное дыхание. Гортань и, в частности, дыхательная щель, – самое узкое место в ВДП, на уровне которого путем рефлекторного изменения расстояния между голосовыми складками автоматически регулируется количество воздуха, поступающего в легкие. При спокойном дыхании голосовая щель находится в состоянии среднего раскрытия и представляет собой просвет треугольной формы с вершиной, обращенной кпереди, и основанием – кзади. Изменение просвета дыхательной щели осуществляется за счет сокращения и расслабления мышц, прикрепляющихся к голосовым отросткам черпаловидных хрящей. При спокойном дыхании голосовые складки в фазе вдоха несколько расходятся, при выдохе с такой же амплитудой сходятся. По-видимому, этот феномен является общим для всей воздухопроводной дыхательной системы, поскольку аналогичным образом реагируют и бронхи.
Величина дыхательной щели в 3–4 раза превышает необходимую для нормального дыхания в покое. Поэтому при срединном положении голосовой складки, возникающем в результате паралича возвратного нерва, когда голосовая щель уменьшается вдвое, больной в покое не испытывает недостатка в кислороде. Необходим лишь некоторый период адаптации к новому состоянию голосовой щели. Во время произвольного глубокого вдоха, при тяжелой физической работе или при патологической гипоксии голосовые складки на вдохе разведены максимально. Такое их разведение осуществляется благодаря сокращению задних перстнечерпаловидных мышц, импульсы к которым могут поступать из пирамидной системы (произвольное сокращение) либо рефлекторно из бульбарного дыхательного центра. В конечном счете дыхательные экскурсии гортани интегрируются физическим актом дыхания как общей моторной функцией дыхательной системы, в основе которой лежит потребность организма в кислороде.
Защитная функция заключается в том, что гортань защищает нижние дыхательные пути от попадания в них инородных тел, жидкости и паров агрессивных химических веществ. Эта функция обеспечивается констрикторами гортани, рефлекторно реагирующими на раздражение ее рецепторов. Физиологический смысл гортанного сфинктера заключается в том, что во время акта глотания, наряду с остановкой дыхания на фазе полувдоха, гортань перекрывается на время прохождения пищевого комка или глотка жидкости по надгортаннику. После соскальзывания пищевого комка, представляющего собой в информационном плане опознанный адекватный объект проглатывания, гортань автоматически раскрывается, вслед за чем неизменно следует рефлекторный выдох, препятствующий аспирации остатков пищи и жидкости, которые могли остаться в гортаноглотке. Сигналом к раскрытию гортани служит прекращение контакта рецепторов надгортанника и слизистой оболочки гортаноглотки с пищевым комком, прошедшим в пищевод. Процесс пересечения пищепроводных и дыхательных путей, происходящий в области гортаноглотки при акте глотания, В. И. Воячек образно назвал механизмом «железнодорожной стрелки».
В отличие от проглатывания естественных продуктов, инородные тела, едкие или горячие жидкости, пары агрессивных веществ распознаются рецепторами гортаноглотки и гортани как объекты угрозы, в результате чего возникает длительный рефлекторный спазм гортани, предохраняющий НДП от попадания в них этих объектов. Однако спазм сам несет в себе угрозу асфиксии. Рефлекторный путь, по которому осуществляется спазм, начинается в чувствительных рецепторах слизистой оболочки нижнего отдела глотки и преддверия гортани. Переключение афферентного пути этого рефлекса на эфферентный происходит в бульбарных моторных центрах, обеспечивающих двигательными нервами констрикторы гортани, диафрагму, межреберные нервы и мышцы брюшного пресса. Поэтому одновременно с сокращением констрикторов гортани, вызывающих закрытие голосовой щели, возникает сокращение указанных мышц и резкое повышение внутригрудного давления с последующим импульсным выдохом (кашлевой рефлекс), в результате чего инородное тело или попавшая в гортань жидкость выталкиваются струей воздуха наружу за пределы гортани. Во время спазма гортани фаза вдоха обычно сопровождается своеобразным стридорозным призвуком.
Мышцы, обеспечивающие защитную функцию гортани, представлены наружными волокнами щиточерпаловидных мышц, межчерпаловидными поперечными и косыми, а также черпалонадгортанными мышцами. При их сокращении происходит сокращение и боковых перстнечерпаловидных мышц. Таким образом, в констрикторной функции гортани участвует много внутренних мышц, способных вызвать мощный спазм голосовой щели, что нередко приводит к фатальным последствиям даже в отсутствие инородного тела, когда причиной спазма могут служить различные нейрогенные факторы.
Кашлевая и отхаркивающая функции не принадлежат собственно гортани, а являются результатом деятельности всего двигательного аппарата дыхательной системы. Кашлевый рефлекс может исходить из чувствительных рецепторов гортани, трахеи, бронхов и плевры. Гортани же принадлежит запирательная функция, обеспечивающая накопление энергии в виде повышения внутригрудного давления с последующим кашлевым толчком при внезапном раскрытии голосовой щели, через которую на большой скорости выдыхаемая воздушная струя увлекает за собой мокроту, пылевые частицы, жидкость, попавшую во время неудачного глотка в гортань и трахею, и т. п.
Кашлевый рефлекс имеет ряд фаз: глубокий вдох (иногда при резком раздражении рецепторов гортани кашлевый рефлекс осуществляется мгновенно на резервном воздухе с последующим стридорозным вдохом), закрытие дыхательной щели, повышение внутригрудного давления в результате сокращения дыхательной мускулатуры, быстрое раскрытие дыхательной щели и взрывное выбрасывание воздуха из бронхов и трахеи. Наиболее чувствительной рефлексогенной зоной является слизистая оболочка межчерпаловидного пространства и бифуркации трахеи. Отсутствие гортани или наличие трахеостомы лишает больного естественного кашлевого рефлекса, что затрудняет эвакуацию мокроты из трахеи и бронхов.
Функция «фиксации» плечевого пояса. В большинстве отечественных учебников и руководств эта функция не упоминается, поскольку она относится к запирательной функции гортани, ее роль в обеспечении максимальных усилий человека, связанных с резким увеличением внутригрудного и внутрибрюшного давления, не освещена вовсе. Тем не менее без этой функции были бы невозможны максимальные усилия, развиваемые мышцами верхнего плечевого пояса, брюшного пресса, мышцами спины и ног. Специальные исследования биомеханики максимальных силовых движений (поднятие тяжестей, спортивная борьба, гребля, метание в легкой атлетике) указывают на то, что в период максимального мышечного усилия верхние конечности активно опираются на грудную клетку посредством плечелопаточной опорно-двигательной системы. Эта опора необходима для максимального использования потенциала усилия, производимого плечевым поясом. Для эффективной реализации этого потенциала грудная клетка должна приобретать ригидность и неподвижность монолита, что достигается глубоким вдохом и полным закрытием дыхательной щели. Во время совершения чрезмерного силового действия, например поднятия рекордного веса, все дыхательные мышцы находятся в максимальном напряжении, что резко повышает внутригрудное и внутрибрюшное давление. Это давление придает грудной клетке так называемую твердость стана, что и позволяет без динамических потерь эффективно использовать потенциальную энергию работающих мышц. Во время реализации максимального усилия, чаще всего в его конечной фазе, из-за чрезмерного внутригрудного давления воздух прорывается через дыхательную щель – и спортсмен издает полунепроизвольный «крик победителя».
Дыхательная щель непроизвольно запирается при необходимости натуживания (роды, дефекация). Отсутствие этой функции у трахеотомированных больных или при недостаточности запирательной функции гортани нейрогенного или анатомического характера (паралич запирательных мышц; у больных после хордэктомии) резко снижает физические возможности лиц (опыты с кистевой и становой динамометрией). Показано, что при равных динамометрических показателях кистевого усилия у здоровых лиц и хронических канюленосителей становая сила у последних была ниже на 20–30 %.
Функция, способствующая кровообращению и оксигенации крови, также редко упоминается в учебниках и руководствах по оториноларингологии. Эта функция незаметна у здоровых лиц при обычной жизнедеятельности, но она играет существенную роль в повышении эффективности оксигенации крови при длительных физических нагрузках и у больных с выраженной гипоксией. Дыхательные экскурсии грудной клетки сопровождаются синхронными колебаниями в ней барометрического давления. Эти колебания передаются на кровеносные сосуды легких, обеспечивая насосную функцию, заключающуюся в наполнении их кровью в фазе вдоха (понижение давления в грудной клетке) и выталкивании крови из сосудов при выдохе (повышение давления). Кроме того, повышение давления в легочной ткани способствует более эффективному растворению кислорода в крови и более эффективной адсорбции углекислого газа. Этот феномен можно проиллюстрировать примером, когда при одышке любого происхождения выдох, производимый через сжатые губы, облегчает ощущение недостаточности воздуха.
Голосовая функция. Вопрос о механизме голосообразования дискутируется на протяжении последних двух веков. Исследования в области физиологии голосового аппарата позволили ответить на вопрос, как колеблются голосовые складки; значительно продвинута проблема генерации их колебаний, однако вопрос, что колеблет голосовые складки, нельзя считать окончательно решенным, поскольку все еще не исследованы многие центральные механизмы регуляции голосовой функции. Не ясно, являются ли колебания голосовых складок процессом активным, задаваемым специальным «генератором», или это процесс пассивный, наподобие тех колебаний, которые генерируют звук в духовых музыкальных инструментах, например в трубе.
Энергоносителем звука, издаваемого этим музыкальным инструментом, является давление воздуха в грудной клетке и в преддверии рта, полость которого может служить аналогом подскладочного пространства. Резонатором служат стенки трубы и столб воздуха, находящийся в трубе, колеблющийся с частотой вибрации губ трубача. При данном механизме звукообразования губы лишь настраиваются на определенную частоту колебаний, однако само колебание является пассивным, обеспечиваемым энергией внутригрудного давления. Тональность звука зависит от статического напряжения губ (от их миоэластических свойств), сила звука – от величины барометрического давления, создаваемого в воздухоносных путях музыканта. Рассматривая принцип пассивной генерации звука на приведенном примере, можно заключить, что образование звука в трубе (гортани) основано на четырех компонентах: силе, колеблющей звукообразующий элемент (губы, голосовые складки); носителе энергии звуковых колебаний (столб воздуха в трубе и дыхательных путях); функции звукообразующего элемента, обладающего собственным адаптационным аппаратом (натяжение и расслабление губ и голосовых складок); акустических свойствах системы резонаторов (столб воздуха в трубе и дыхательных путях, стенки трубы, биорезонаторы дыхательных путей и головы).
Все указанные компоненты звукообразования, на первый взгляд, вполне удовлетворительно укладываются в сущность так называемой миоэластической теории голосообразования, разработка которой началась за 15 лет до изобретения М. Гарсией непрямой ларингоскопии. Экспериментальную основу науки о голосе человека заложил И. Мюллер[4] (1839), сконструировавший искусственную гортань, в которой роль голосовых складок играли резиновые полоски, а воздушный напор создавался мехами. На основании полученных данных автор стал рассматривать гортань как подобие язычкового музыкального инструмента, вибраторы которого приходят в пассивные колебания благодаря энергии воздушного потока. Эта концепция господствовала вплоть до середины XX в., когда появились труды французского физиолога Р. Юссона, показавшего, что колебания голосовых складок – отнюдь не пассивное явление, оно имеет в своей основе нейрохронаксические процессы, обеспечивающие центральную нейромоторную функцию голосовых складок.
Сущность миоэластической теории И. Мюллера, которая не утратила актуальности, заключается в следующем. Перед звукообразованием дыхательный цикл прекращается на полувдохе, голосовые складки под влиянием сократительной функции приводящих мышц занимают срединное положение, замыкая голосовую щель, и приобретают необходимые для данного тона напряжение, толщину и длину, т. е. настраиваются на определенную собственную частоту колебаний. Одновременно с замыканием голосовой щели под влиянием сокращения дыхательных мышц повышается барометрическое давление в легких, бронхах, трахее и подскладочном пространстве. Местом наименьшего сопротивления этому воздушному подпору являются сомкнутые голосовые складки. Когда давление преодолевает силу сомкнутых голосовых складок, воздух прорывается через голосовую щель, в результате чего давление в подскладочном пространстве падает и голосовые складки вновь смыкаются. Это приводит к новому увеличению давления в подскладочном пространстве – и цикл многократно повторяется с частотой издаваемого звука. Частота звука зависит от степени смыкания голосовых складок, их длины и толщины, а сила звука – от сократительного усилия аддукторов голосовых складок и величины подскладочного барометрического давления. Указанный процесс характеризуется постоянным противоборством между подскладочным давлением и упругими силами, смыкающими голосовые складки (теория «голосовой борьбы»). Таким образом, главным тезисом миоэластической теории является то, что колебания голосовых складок при голосообразовании пассивны и в их основе лежит чисто механический процесс. Однако, как показали последующие исследования, с помощью миоэластической теории нельзя объяснить ряд физиологических явлений; кроме того, в самой теории имеются существенные противоречия.
Образование низких звуков обеспечивается только mm. vocales, которые максимально напрягаются и утолщаются. В таком состоянии они способны колебаться только в узком низкочастотном диапазоне, при этом их натянутость минимальна, как у отпущенной струны. Расстояние между щитовидным и перстневидным хрящами максимальное, что приводит к увеличению резонирующего объема гортани, настраивающегося на низкие частоты. Как полагал В. Г. Ермолаев[5] (1958), такой механизм голосообразования используется для воспроизведения звуков нижней четверти голосового диапазона, присущего данному человеку.
Рис. 18.6. Нервы гортани, вид слева:
1 – возвратный нерв; 2 – нижний сегмент петли Галена[3]; 3, 6 – волокна к щиточерпаловидной мышце; 4 – латеральная перстнечерпаловидная мышца; 5 – петля Галена; 7 – черпаловидный хрящ; 8 – продолжение верхнего гортанного нерва; 9 – наружная ветвь верхнего гортанного нерва; 10 – ствол верхнего гортанного нерва; 11 – нервные волокна к надгортаннику; 12, 13 – косые и поперечные волокна черпало-черпаловидной мышцы; 14 – нервные волокна к черпало-черпаловидной мышце; 15 – задняя перстнечерпаловидная мышца
Дыхательная функция гортани заключается не в пассивном пропускании воздуха в легкие и обратно, а в ряде активных рефлекторных актов, совершаемых в гортани и оптимизирующих легочное дыхание. Гортань и, в частности, дыхательная щель, – самое узкое место в ВДП, на уровне которого путем рефлекторного изменения расстояния между голосовыми складками автоматически регулируется количество воздуха, поступающего в легкие. При спокойном дыхании голосовая щель находится в состоянии среднего раскрытия и представляет собой просвет треугольной формы с вершиной, обращенной кпереди, и основанием – кзади. Изменение просвета дыхательной щели осуществляется за счет сокращения и расслабления мышц, прикрепляющихся к голосовым отросткам черпаловидных хрящей. При спокойном дыхании голосовые складки в фазе вдоха несколько расходятся, при выдохе с такой же амплитудой сходятся. По-видимому, этот феномен является общим для всей воздухопроводной дыхательной системы, поскольку аналогичным образом реагируют и бронхи.
Величина дыхательной щели в 3–4 раза превышает необходимую для нормального дыхания в покое. Поэтому при срединном положении голосовой складки, возникающем в результате паралича возвратного нерва, когда голосовая щель уменьшается вдвое, больной в покое не испытывает недостатка в кислороде. Необходим лишь некоторый период адаптации к новому состоянию голосовой щели. Во время произвольного глубокого вдоха, при тяжелой физической работе или при патологической гипоксии голосовые складки на вдохе разведены максимально. Такое их разведение осуществляется благодаря сокращению задних перстнечерпаловидных мышц, импульсы к которым могут поступать из пирамидной системы (произвольное сокращение) либо рефлекторно из бульбарного дыхательного центра. В конечном счете дыхательные экскурсии гортани интегрируются физическим актом дыхания как общей моторной функцией дыхательной системы, в основе которой лежит потребность организма в кислороде.
Защитная функция заключается в том, что гортань защищает нижние дыхательные пути от попадания в них инородных тел, жидкости и паров агрессивных химических веществ. Эта функция обеспечивается констрикторами гортани, рефлекторно реагирующими на раздражение ее рецепторов. Физиологический смысл гортанного сфинктера заключается в том, что во время акта глотания, наряду с остановкой дыхания на фазе полувдоха, гортань перекрывается на время прохождения пищевого комка или глотка жидкости по надгортаннику. После соскальзывания пищевого комка, представляющего собой в информационном плане опознанный адекватный объект проглатывания, гортань автоматически раскрывается, вслед за чем неизменно следует рефлекторный выдох, препятствующий аспирации остатков пищи и жидкости, которые могли остаться в гортаноглотке. Сигналом к раскрытию гортани служит прекращение контакта рецепторов надгортанника и слизистой оболочки гортаноглотки с пищевым комком, прошедшим в пищевод. Процесс пересечения пищепроводных и дыхательных путей, происходящий в области гортаноглотки при акте глотания, В. И. Воячек образно назвал механизмом «железнодорожной стрелки».
В отличие от проглатывания естественных продуктов, инородные тела, едкие или горячие жидкости, пары агрессивных веществ распознаются рецепторами гортаноглотки и гортани как объекты угрозы, в результате чего возникает длительный рефлекторный спазм гортани, предохраняющий НДП от попадания в них этих объектов. Однако спазм сам несет в себе угрозу асфиксии. Рефлекторный путь, по которому осуществляется спазм, начинается в чувствительных рецепторах слизистой оболочки нижнего отдела глотки и преддверия гортани. Переключение афферентного пути этого рефлекса на эфферентный происходит в бульбарных моторных центрах, обеспечивающих двигательными нервами констрикторы гортани, диафрагму, межреберные нервы и мышцы брюшного пресса. Поэтому одновременно с сокращением констрикторов гортани, вызывающих закрытие голосовой щели, возникает сокращение указанных мышц и резкое повышение внутригрудного давления с последующим импульсным выдохом (кашлевой рефлекс), в результате чего инородное тело или попавшая в гортань жидкость выталкиваются струей воздуха наружу за пределы гортани. Во время спазма гортани фаза вдоха обычно сопровождается своеобразным стридорозным призвуком.
Мышцы, обеспечивающие защитную функцию гортани, представлены наружными волокнами щиточерпаловидных мышц, межчерпаловидными поперечными и косыми, а также черпалонадгортанными мышцами. При их сокращении происходит сокращение и боковых перстнечерпаловидных мышц. Таким образом, в констрикторной функции гортани участвует много внутренних мышц, способных вызвать мощный спазм голосовой щели, что нередко приводит к фатальным последствиям даже в отсутствие инородного тела, когда причиной спазма могут служить различные нейрогенные факторы.
Кашлевая и отхаркивающая функции не принадлежат собственно гортани, а являются результатом деятельности всего двигательного аппарата дыхательной системы. Кашлевый рефлекс может исходить из чувствительных рецепторов гортани, трахеи, бронхов и плевры. Гортани же принадлежит запирательная функция, обеспечивающая накопление энергии в виде повышения внутригрудного давления с последующим кашлевым толчком при внезапном раскрытии голосовой щели, через которую на большой скорости выдыхаемая воздушная струя увлекает за собой мокроту, пылевые частицы, жидкость, попавшую во время неудачного глотка в гортань и трахею, и т. п.
Кашлевый рефлекс имеет ряд фаз: глубокий вдох (иногда при резком раздражении рецепторов гортани кашлевый рефлекс осуществляется мгновенно на резервном воздухе с последующим стридорозным вдохом), закрытие дыхательной щели, повышение внутригрудного давления в результате сокращения дыхательной мускулатуры, быстрое раскрытие дыхательной щели и взрывное выбрасывание воздуха из бронхов и трахеи. Наиболее чувствительной рефлексогенной зоной является слизистая оболочка межчерпаловидного пространства и бифуркации трахеи. Отсутствие гортани или наличие трахеостомы лишает больного естественного кашлевого рефлекса, что затрудняет эвакуацию мокроты из трахеи и бронхов.
Функция «фиксации» плечевого пояса. В большинстве отечественных учебников и руководств эта функция не упоминается, поскольку она относится к запирательной функции гортани, ее роль в обеспечении максимальных усилий человека, связанных с резким увеличением внутригрудного и внутрибрюшного давления, не освещена вовсе. Тем не менее без этой функции были бы невозможны максимальные усилия, развиваемые мышцами верхнего плечевого пояса, брюшного пресса, мышцами спины и ног. Специальные исследования биомеханики максимальных силовых движений (поднятие тяжестей, спортивная борьба, гребля, метание в легкой атлетике) указывают на то, что в период максимального мышечного усилия верхние конечности активно опираются на грудную клетку посредством плечелопаточной опорно-двигательной системы. Эта опора необходима для максимального использования потенциала усилия, производимого плечевым поясом. Для эффективной реализации этого потенциала грудная клетка должна приобретать ригидность и неподвижность монолита, что достигается глубоким вдохом и полным закрытием дыхательной щели. Во время совершения чрезмерного силового действия, например поднятия рекордного веса, все дыхательные мышцы находятся в максимальном напряжении, что резко повышает внутригрудное и внутрибрюшное давление. Это давление придает грудной клетке так называемую твердость стана, что и позволяет без динамических потерь эффективно использовать потенциальную энергию работающих мышц. Во время реализации максимального усилия, чаще всего в его конечной фазе, из-за чрезмерного внутригрудного давления воздух прорывается через дыхательную щель – и спортсмен издает полунепроизвольный «крик победителя».
Дыхательная щель непроизвольно запирается при необходимости натуживания (роды, дефекация). Отсутствие этой функции у трахеотомированных больных или при недостаточности запирательной функции гортани нейрогенного или анатомического характера (паралич запирательных мышц; у больных после хордэктомии) резко снижает физические возможности лиц (опыты с кистевой и становой динамометрией). Показано, что при равных динамометрических показателях кистевого усилия у здоровых лиц и хронических канюленосителей становая сила у последних была ниже на 20–30 %.
Функция, способствующая кровообращению и оксигенации крови, также редко упоминается в учебниках и руководствах по оториноларингологии. Эта функция незаметна у здоровых лиц при обычной жизнедеятельности, но она играет существенную роль в повышении эффективности оксигенации крови при длительных физических нагрузках и у больных с выраженной гипоксией. Дыхательные экскурсии грудной клетки сопровождаются синхронными колебаниями в ней барометрического давления. Эти колебания передаются на кровеносные сосуды легких, обеспечивая насосную функцию, заключающуюся в наполнении их кровью в фазе вдоха (понижение давления в грудной клетке) и выталкивании крови из сосудов при выдохе (повышение давления). Кроме того, повышение давления в легочной ткани способствует более эффективному растворению кислорода в крови и более эффективной адсорбции углекислого газа. Этот феномен можно проиллюстрировать примером, когда при одышке любого происхождения выдох, производимый через сжатые губы, облегчает ощущение недостаточности воздуха.
Голосовая функция. Вопрос о механизме голосообразования дискутируется на протяжении последних двух веков. Исследования в области физиологии голосового аппарата позволили ответить на вопрос, как колеблются голосовые складки; значительно продвинута проблема генерации их колебаний, однако вопрос, что колеблет голосовые складки, нельзя считать окончательно решенным, поскольку все еще не исследованы многие центральные механизмы регуляции голосовой функции. Не ясно, являются ли колебания голосовых складок процессом активным, задаваемым специальным «генератором», или это процесс пассивный, наподобие тех колебаний, которые генерируют звук в духовых музыкальных инструментах, например в трубе.
Энергоносителем звука, издаваемого этим музыкальным инструментом, является давление воздуха в грудной клетке и в преддверии рта, полость которого может служить аналогом подскладочного пространства. Резонатором служат стенки трубы и столб воздуха, находящийся в трубе, колеблющийся с частотой вибрации губ трубача. При данном механизме звукообразования губы лишь настраиваются на определенную частоту колебаний, однако само колебание является пассивным, обеспечиваемым энергией внутригрудного давления. Тональность звука зависит от статического напряжения губ (от их миоэластических свойств), сила звука – от величины барометрического давления, создаваемого в воздухоносных путях музыканта. Рассматривая принцип пассивной генерации звука на приведенном примере, можно заключить, что образование звука в трубе (гортани) основано на четырех компонентах: силе, колеблющей звукообразующий элемент (губы, голосовые складки); носителе энергии звуковых колебаний (столб воздуха в трубе и дыхательных путях); функции звукообразующего элемента, обладающего собственным адаптационным аппаратом (натяжение и расслабление губ и голосовых складок); акустических свойствах системы резонаторов (столб воздуха в трубе и дыхательных путях, стенки трубы, биорезонаторы дыхательных путей и головы).
Все указанные компоненты звукообразования, на первый взгляд, вполне удовлетворительно укладываются в сущность так называемой миоэластической теории голосообразования, разработка которой началась за 15 лет до изобретения М. Гарсией непрямой ларингоскопии. Экспериментальную основу науки о голосе человека заложил И. Мюллер[4] (1839), сконструировавший искусственную гортань, в которой роль голосовых складок играли резиновые полоски, а воздушный напор создавался мехами. На основании полученных данных автор стал рассматривать гортань как подобие язычкового музыкального инструмента, вибраторы которого приходят в пассивные колебания благодаря энергии воздушного потока. Эта концепция господствовала вплоть до середины XX в., когда появились труды французского физиолога Р. Юссона, показавшего, что колебания голосовых складок – отнюдь не пассивное явление, оно имеет в своей основе нейрохронаксические процессы, обеспечивающие центральную нейромоторную функцию голосовых складок.
Сущность миоэластической теории И. Мюллера, которая не утратила актуальности, заключается в следующем. Перед звукообразованием дыхательный цикл прекращается на полувдохе, голосовые складки под влиянием сократительной функции приводящих мышц занимают срединное положение, замыкая голосовую щель, и приобретают необходимые для данного тона напряжение, толщину и длину, т. е. настраиваются на определенную собственную частоту колебаний. Одновременно с замыканием голосовой щели под влиянием сокращения дыхательных мышц повышается барометрическое давление в легких, бронхах, трахее и подскладочном пространстве. Местом наименьшего сопротивления этому воздушному подпору являются сомкнутые голосовые складки. Когда давление преодолевает силу сомкнутых голосовых складок, воздух прорывается через голосовую щель, в результате чего давление в подскладочном пространстве падает и голосовые складки вновь смыкаются. Это приводит к новому увеличению давления в подскладочном пространстве – и цикл многократно повторяется с частотой издаваемого звука. Частота звука зависит от степени смыкания голосовых складок, их длины и толщины, а сила звука – от сократительного усилия аддукторов голосовых складок и величины подскладочного барометрического давления. Указанный процесс характеризуется постоянным противоборством между подскладочным давлением и упругими силами, смыкающими голосовые складки (теория «голосовой борьбы»). Таким образом, главным тезисом миоэластической теории является то, что колебания голосовых складок при голосообразовании пассивны и в их основе лежит чисто механический процесс. Однако, как показали последующие исследования, с помощью миоэластической теории нельзя объяснить ряд физиологических явлений; кроме того, в самой теории имеются существенные противоречия.
Образование низких звуков обеспечивается только mm. vocales, которые максимально напрягаются и утолщаются. В таком состоянии они способны колебаться только в узком низкочастотном диапазоне, при этом их натянутость минимальна, как у отпущенной струны. Расстояние между щитовидным и перстневидным хрящами максимальное, что приводит к увеличению резонирующего объема гортани, настраивающегося на низкие частоты. Как полагал В. Г. Ермолаев[5] (1958), такой механизм голосообразования используется для воспроизведения звуков нижней четверти голосового диапазона, присущего данному человеку.