Страница:
Гендерная идентичность как биологический процесс: нормальная пренатальная дифференциация
С момента зачатия на дифференциацию полов на мужской и женский оказывают влияние многочисленные биологические факторы. На следующих страницах мы проанализируем, как происходит биологическая половая дифференциация в период пренатального развития. Наше описание будет построено по хронологическому принципу: мы начнем с момента зачатия и рассмотрим различия мужских и женских хромосом. Затем мы перейдем к развитию гонад, выработке гормонов, формированию внутренней и наружной репродуктивных структур и закончим рассмотрением половой дифференциации мозга.
Пол на хромосомном уровне
Наш биологический пол определяется при зачатии хромосомным составом сперматозоида(мужской половой клетки), оплодотворяющего яйцеклетку, или яйцо (женскую половую клетку). Все клетки человеческого организма, за исключением половых клеток, содержат набор из 46 хромосом, состоящий из 23 пар (рис. 3.1). Двадцать две из них являются комплементарными. Иными словами, хромосомы, входящие в состав каждой пары, практически идентичны. Эти комплементарные пары, называемые автосомами, одинаковы и у мужчин и у женщин. Они не оказывают влияния на дифференциацию полов. Однако одна хромосомная пара — пара половых хромосом— у мужчин и женщин различная. Женщины имеют две сходные хромосомы, обозначаемые XX, тогда как мужчины имеют различные хромосомы, обозначаемые XY.
Сперматозоид. Мужская половая клетка.
Яйцеклетка. Женская половая клетка.
Автосомы. 22 пары человеческих хромосом, не оказывающих значительного влияния на дифференциацию полов.
Половые хромосомы. Единичный набор хромосом, оказывающий определяющее влияние на биологический пол.
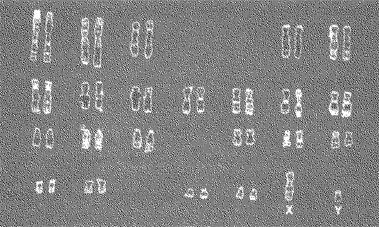
Рис. 3.1. Человеческая клетка содержит 22 пары комплементарных (идентичных друг другу) автосом и одну пару половых хромосом. В норме женщина имеет две X-хромосомы, а мужчина — X-хромосому и Y-хромосому
Как уже отмечалось выше, половые клетки являются исключением из правила, распространяющегося на весь 23-парный набор. В результате биологического процесса, известного как мейоз, созревшие половые клетки содержат только половину полного набора хромосом — по одному компоненту каждой пары. (Данный процесс необходим, чтобы избежать удвоения полного числа хромосом, когда половые клетки соединяются при зачатии.) Нормальная женская яйцеклетка (или яйцо) содержит 22 автосомы плюс одну X-хромосому. Нормальный мужской сперматозоид содержит 22 автосомы плюс либо X-, либо Y-хромосому. В яйцеклетке, оплодотворенной сперматозоидом, несущим Y-хромосому, образуется комбинация XY. В результате этого рождается мальчик. С другой стороны, если яйцеклетку оплодотворяет Х-несущий сперматозоид, образуется комбинация XX. В результате этого рождается девочка. Для формирования полноценных внутренних и наружных структур женского организма необходимы две X-хромосомы. Если же в наборе присутствует одна Y-хромосома, у ребенка формируются мужские наружные половые органы и органы размножения (Harley et al., 1992; Page et al., 1987).
Недавно исследователям удалось локализовать единичный ген на короткой ветви человеческой Y-хромосомы. Этот ген, по-видимому, и играет решающую роль в запуске последовательности событий, приводящих к формированию мужских гонад, или яичек. (Как мы вскоре увидим, яички, в свою очередь, выделяют гормоны, стимулирующие формирование других структур мужского организма.) «Мужской» ген, ответственный за формирование мужского организма (определяющий появление яичек), получил название SRY-гена (Jegalian & Lahn, 2001; Terrinoni et al., 2000).
Яички. Мужские гонады, расположенные в мошонке и вырабатывающие сперму и половые гормоны.
Результаты исследования, проведенного итальянскими и американскими учеными, позволяют предположить, что, возможно, существует также и «женский» ген или гены, ответственные за формирование женского организма. Ученые исследовали четыре случая обратного превращения мужских организмов в женские у индивидов, обладающих XY-хромосомами и функционирующими SRY (мужскими) генами.
У троих из четырех исследуемых лиц абсолютно отчетливо наблюдались женские внешние половые органы. Половые органы четвертого индивида носили амбивалентный характер. Если бы мужской ген являлся решающим фактором, определяющим биологический пол, подобных обратных превращений не могло бы произойти. Что же явилось причиной этих превращений? В ходе анализа ДНК исследуемых лиц был обнаружен факт дупликации крохотного участка генетического материала на короткой ветви X-хромосом. В результате, каждый из этих индивидов обладал двойной дозой гена, получившего обозначение DSS. Именно этот факт и явился причиной превращения в остальном нормального мужского плода в женский (Bardoni et al., 1994).
Полученные результаты позволяют предположить, что ген (или гены), содержащиеся в X-хромосоме, способствуют развитию недифференцированных гонад в женском направлении, аналогично тому как SRY-ген способствует началу формирования мужских половых структур. Данное наблюдение противоречит господствовавшему в течение длительного времени представлению о том, что человеческий зародыш является изначально женским. Противоречит оно и убеждению, что в отличие от пренатальной дифференциации, ведущей к появлению мужского организма, для дифференциации, ведущей к появлению женского организма, не требуется наличия запускающих факторов на генетическом уровне.
Пол на гонадном уровне
В первые недели после зачатия структуры позднее развивающиеся в органы размножения, или гонады, одинаковы у мужчин и женщин (рис. 3.2, а). Дифференциация начинается спустя приблизительно 6 недель после зачатия. Разовьется ли масса недифференцированных половых тканей в мужские или женские гонады, определяется генетическими факторами (Clarnette et al., 1997). К этому моменту продукт (или продукты) SRY-гена мужского плода инициируют процесс превращения зародышевых гонад в яички. Если продукты SRY-гена отсутствуют, а также, вероятно, под влиянием DSS или других женских генов, недифференцированные гонадные ткани развиваются в яичники(рис. 3.2, б).
Гонады. Мужские и женские половые железы — яички и яичники.
Яичники. Женские гонады, производящие яйцеклетки и половые гормоны.
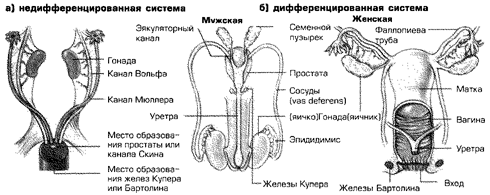
По окончании процесса формирования яичники или яички начинают выделять собственные половые гормоны. Как мы увидим далее, эти гормоны становятся решающим фактором в дальнейшей половой дифференциации. Генетические же влияния прекращают свое действие.
Пол на гормональном уровне
Как и другие железы эндокринной системы(системы бесканальных желез, включающих гипофиз, щитовидную, паращитовидную, надпочечниковые и поджелудочную железы), гонады вырабатывают гормоны и выделяют их непосредственно в кровь. Яичники вырабатывают два типа гормонов: эстрогеныи прогестационные соединения. Эстрогены, наиболее важным из которых является эстрадиол, оказывают влияние на развитие женских физических признаков и участвуют в регулировании менструального цикла. Наиболее важным с физиологической точки зрения из известных прогестационных соединений является прогестерон. Его функции включают участие в регулировании менструального цикла и стимуляцию развития стенок матки в периоды подготовки к беременности. Основными гормональными продуктами яичек являются андрогены. Наиболее важный из них, тестостерон, оказывает влияние на развитие мужских физических половых признаков и сексуальной мотивации. У обоих полов надпочечниковые железы также выделяют половые гормоны, включая небольшое количество эстрогена и большое количество андрогена.
Эстрогены. Класс гормонов, регулирующих менструальный цикл и ответственных за развитие вторичных женских половых признаков.
Прогестационные соединения. Класс гормонов, включая прогестерон, вырабатываемый яичниками.
Андрогены. Класс гормонов, способствующих развитию мужских гениталий и вторичных половых признаков, а также оказывающих влияние на сексуальную мотивацию обоих полов. Данные гормоны вырабатываются надпочечниковыми железами как у мужчин, так и у женщин, а также яичками у мужчин.
Пол на уровне внутренних репродукционных структур
Приблизительно на 8-й неделе после зачатия половые гормоны начинают играть важную роль в дифференциации полов. К этому времени две проводящие системы, показанные на рис. 3.2, a — каналы Вольфаи каналы Мюллера— начинают дифференцироваться, формируясь во внутренние структуры, показанные на рис. 3.2, б. У мужского плода андрогены, выделяемые яичками, стимулируют развитие каналов Вольфа и превращение их в семявыносящие протоки, семенные пузырьки и эякуляторный канал. Другое выделяемое яичками вещество известно как тормозящее вещество Мюллера (Mullerian inhibiting substance, MIS). Это вещество вызывает усыхание и исчезновение системы каналов Мюллера у мужчин (Clarence et al., 1997; Lee et al., 1997). Каналы Мюллера развиваются в фаллопиевы трубы, матку и внутреннюю треть вагины, а проводящая система Вольфа атрофируется.
Пол на уровне наружных половых органов (гениталий)
Наружные половые органы формируются аналогичным образом. До тех пор пока гонады не начнут выделять собственные гормоны на 6-й неделе после зачатия, ткани мужских и женских наружных половых органов остаются недифференцированными (рис. 3.3). Из этих тканей начинают формироваться либо мужские, либо женские гениталии. Это зависит от присутствия либо отсутствия продукта тестостерона, выделяемого у мужчин и известного как дигидротестостерон (ДГТ). ДГТ стимулирует развитие мошонки из мошоночной выпуклости. Из генитальных туберкули генитальных складокначинают развиваться гланы (головки) и ствол пениса соответственно. Генитальные складки срастаются вокруг уретры, формируя ствол пениса, а оба края мошоночной выпуклости срастаются, формируя мошонку. У женщин подобного срастания не происходит. Если отсутствует тестостерон (а также, вероятно, под влиянием веществ, стимулируемых DSS или женским геном), генитальные туберкулы превращаются в клитор, а генитальные складки — в малые половые губы. Оба края мошоночной выпуклости при этом дифференцируются в большие половые губы. К 12-й неделе процесс дифференциации завершается: у плода мужского рода отчетливо видны пенис и мошонка, а у плода женского рода — половые губы и клитор.
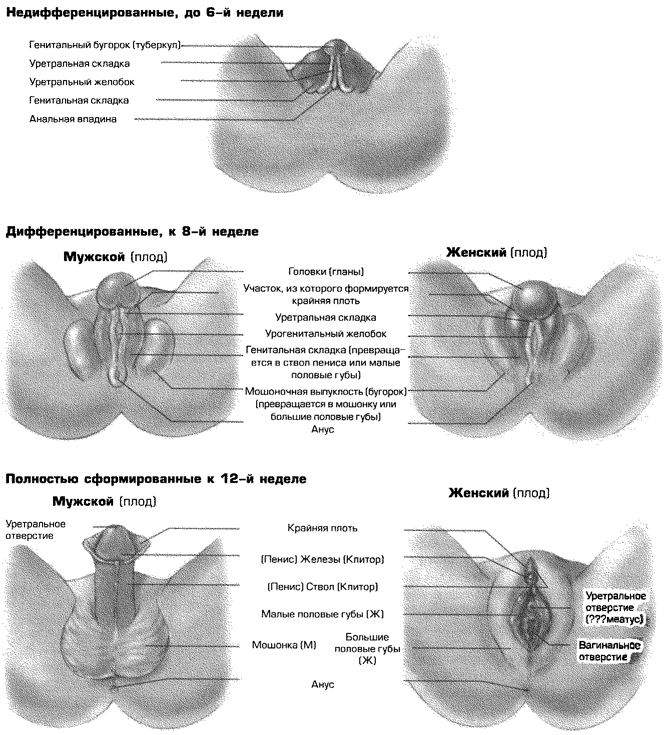
Рис. 3.3. Пренатальное развитие мужских и женских наружных половых органов из недифференцированных в дифференцированные
Поскольку наружные половые органы, гонады, а также ряд внутренних структур у мужчин и женщин развиваются из одних и тех же зародышевых тканей, неудивительно, что они являются соответствующими друг другу, или гомологичными, органами. В табл. 3.1 приводятся соответствия женских и мужских органов.
Таблица 3.1. Гомологичные половые органы

Половая дифференциация мозга
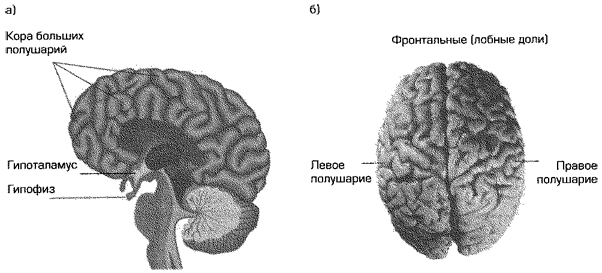
Результаты исследований позволяют предположить, что существует ряд значимых функциональных и структурных различий между мозгом мужчины и женщины. Эти различия являются результатом, по крайней мере отчасти, пренатального процесса половой дифференциации (Gur et al., 1995; Reiner, 1997; Witelson, 1991). Они касаются как минимум двух основных отделов мозга: гипоталамуса, а также левого и правого полушарий головного мозга(рис. 3.4).
Рядом исследователей была обнаружена связь между различиями мужского и женского гипоталамуса и присутствием либо отсутствием тестостерона в процессе пренатальной дифференциации (Reiner, 1997; Zhou et al., 1995). При отсутствии циркуляции тестостерона в женском гипоталамусе развиваются специализированные рецепторные клетки, исключительно восприимчивые к присутствию эстрогена в крови. В зародыше мужского организма присутствие тестостерона препятствует развитию у этих клеток восприимчивости к эстрогену. Данная форма пренатальной дифференциации является ключевым фактором, оказывающим влияние на все последующие события. В период полового созревания восприимчивый к эстрогену женский гипоталамус управляет работой гипофиза, побуждая его выделять гормоны согласно циклическому ритму, запуская таким образом менструальный цикл. У мужчин невосприимчивый к эстрогену гипоталамус управляет стабильной (неритмической) выработкой половых гормонов.
Исследователям удалось обнаружить любопытные факты, касающиеся половой дифференциации крохотного участка гипоталамуса, так называемого bed nucleus of the strea terminalis (BST). Ученые продемонстрировали, что BST содержит рецепторы эстрогена и андрогена и оказывает определяющее влияние на сексуальное поведение животных (высших млекопитающих) (Allen & Gorski, 1990; Breedlove, 1995). В отчете об одном недавно проведенном исследовании сообщалось, что центральный отдел BST (BSTc) у гетеросексуальных мужчин на 44 % крупнее, чем у гетеросексуальных женщин (Zhou et al., 1995). Авторы данного исследования выдвинули гипотезу, согласно которой различия в размерах BST у взрослых людей определяются в период пренатального развития и являются функцией организованного воздействия половых гормонов. В ходе другого исследования было обнаружено, что специфический хвостовой, или задний, отдел BST (BNST-dspm) в 2,5 раза крупнее у мужчин, чем у женщин (Allen & Gorski, 1990). Факт обнаружения половых различий в этих двух участках BST, вероятно, свидетельствует о том, что данный отдел гипоталамуса может оказывать влияние на половые различия, а также на сексуальное функционирование людей.
Исследователи также обнаружили половые различия переднего отдела гипоталамуса, называемого преоптической зоной (preoptic area, POA). В ходе двух исследований был локализован специфический участок этой зоны, названный ядро полового диморфизма преоптической зоны (sexually dimorphic nucleus of the preoptic area (SND-POA), значительно более крупный у взрослых мужчин, чем у взрослых женщин (Allen et al., 1989; Swaab & Fliers, 1985). Ученые предположили, что данное различие также обусловлено воздействием половых гормонов в период пренатального развития.
Эти и другие многочисленные исследования, перечисление которых заняло бы слишком много места, наглядно продемонстрировали, что в человеческом гипоталамусе имеют место специфические значимые половые различия. Полученные данные являются веским подтверждением гипотезы о том, что существует биологический субстрат половых различий, касающихся сексуального поведения людей (Diamond & Sigmundson, 1997).
Однако наше понимание истинной природы и механизмов этих различий еще нуждается в дальнейшем прояснении в ходе будущих исследований. Поэтому нам следует проявлять осторожность, приписывая поведенческие половые различия воздействию биологических факторов. Тем не менее растет число свидетельств в пользу влияния пренатальных половых гормонов на внутриутробное развитие мозга. Это позволило некоторым ученым высказать предположение, что специфическое гендерное поведение детей и взрослых по крайней мере отчасти является результатом общей гормонально-обусловленной маскулинизации или феминизации мозга в период пренатального развития (Coolaer & Hines, Durden-Smith & de Simone, 1995; Lerman et al., 2000). С точки зрения этих ученых представляется возможным, что «вероятно, органом, оказывающим решающее влияние на психосексуальное развитие и адаптацию, является мозг, а не наружные половые органы» (Reiner, 1997, р. 225).
Также были обнаружены половые различия в структуре больших полушарий мозга и в степени специализации полушарий, касающейся вербальных и пространственных когнитивных навыков (Diamond, 1991; Shaywitz et al., 1995). Женщины часто получают более высокие оценки по тестам развития вербальных навыков, чем мужчины. Для пространственных же навыков верно обратное (Halpern, 1992; Voyer et al., 1995). Некоторые ученые полагают, что различия в структуре полушарий, возможно, являются биологическим базисом данных проявлений различия между мужчинами и женщинами (Geschwind & Behan, 1984; Witelson, 1988, 1991).
Тем не менее многие ученые считают, что отмечаемые исследователями половые различия, касающиеся когнитивных навыков, в основном определяются психосоциальными факторами (Geary, 1989; Hyde, 1996). Они приводят убедительные свидетельства того, что в последние годы подобные различия начали стремительно исчезать (Hyde & Plant, 1995; Voyer, 1988, 1991).
Результаты двух исследований, проведенных Американской ассоциацией университетских сотрудниц (American Association of University Women, AAUW), представляют собой неоспоримые факты, свидетельствующие о наличии тенденции в сторону когнитивного равенства полов. В 1992 году данная организация опубликовала сообщение о том, что в начале школьного обучения девочки демонстрируют способности к математике и естественным наукам, не уступающие способностям мальчиков. Однако к моменту окончания высшей (восьмилетней) школы девушки значительно отстают от юношей по этим когнитивным показателям (AAUW, 1992). В ходе повторного исследования, проведенного в 1988 году, было обнаружено, что разрыв в уровне достижений между представителями обоего пола в области математики и естественных наук практически исчез, за исключением некоторых специфических разделов высшей математики (Carter, 2000).
Некоторые авторы утверждают, что биологические объяснения половых различий, касающихся когнитивных (и поведенческих) проявлений, отнюдь не имеют под собой серьезного научного основания и выступают лишь в качестве ничем не подкрепленной попытки оправдать сохранение в современном обществе традиционных гендерных ролей. (См., в частности, книгу, написанную специалистом по генетике индивидуального развития Энн Фаусто-Стэрлинг «Половое разделение тела: Гендерная политика и формирование сексуальности» (Anne Fausto-Sterling, Sexing the Body: Gender Politics and the Construction of Sexuality, 1999).)
В прошлом многие представительницы феминистского движения крайне неохотно признавали факт различия полов, утверждая, что все, кроме наиболее очевидных половых различий, является «продуктами культуры, а не генов, и может быть устранено путем введения соответствующего законодательства и методов воспитания детей» (Ehrenreich, 1999, р. 58).
В настоящее время, однако, некоторые авторитетные женщины-ученые не только признают, но и многозначительно подчеркивают факт существования половых различий на церебральном уровне, включая и те из них, которые, вероятно, являются биологическими по своей природе. (См., в частности, книги: Натали Анжье «Женщина: Интимная география» (Natalie Angier, Woman: An Intimate Geography, 1999); Хелен Фишер «Первородный пол: Природные способности женщин, как они изменят мир» (Helen Fisher, The First Sex: The Natural Talents of Women and How They Will Change the World, 1999); Диана Хэйлз «Совсем как женщина: Пересматривая представления о том, что делает нас женщинами, в свете гендерной науки» (Dianne Hales, Just Like a Woman: How Gender Science Is Redefining What Makes Us Female, 1999).) Поскольку поток новых данных постоянно растет, возможно, в ближайшем будущем мы сможем делать обоснованные заключения относительно того, существует ли биологический базис половых различий, касающихся когнитивных навыков и поведенческих черт.
Аномальная пренатальная дифференциация
До сих пор мы рассматривали только нормальную (типичную) пренатальную дифференциацию. Однако значительная часть известных нам данных о влиянии биологической половой дифференциации на формирование гендерной идентичности получена в ходе исследования случаев аномальной (нетипичной) дифференциации.
Мы уже говорили о том, что дифференциация внутренних и наружных половых структур происходит под влиянием биологических факторов (запускающих сигналов). Результатом отклонений механизмов этих сигналов от нормальных может стать амбивалентность биологического пола. Людей, обладающих амбивалентными или противоречивыми половыми признаками, иногда называют гермафродитами. Это термин, заимствованный из греческой мифологии и обозначающий божество Гермафродита, по представлениям древних греков обладавшего атрибутами обоих полов. Сегодня вместо термина «гермафродит» по отношению к таким людям все чаще начинает употребляться термин «интерсексуал».
Интерсексуалы. См. псевдогермафродиты.
Мы можем различить истинных гермафродитови псевдогермафродитов. Истинные гермафродиты, в организме которых присутствуют ткани как яичников, так и яичек, встречаются крайне редко (Parker, 1998; Unlu et al., 1997). Их наружные половые органы часто представляют собой сочетание женских и мужских структур. Псевдогермафродиты встречаются значительно чаще, что в процентном отношении составляет приблизительно 1:2000 (Colapinto, 2000). Эти индивиды также отличаются амбивалентной внутренней и наружной репродуктивной анатомией. Однако в отличие от истинных гермафродитов псевдогермафродиты рождаются с гонадами, соответствующими их хромосомному полу. Исследования псевдогермафродитов позволили прояснить относительную роль биологических факторов и социального научения в формировании гендерной идентичности. Интерсексуальность может иметь место вследствие аномальной комбинации половых хромосом или в результате пренатальных гормональных нарушений. В этом разделе мы рассмотрим данные, полученные в ходе изучения пяти разновидностей псевдогермафродизма. Результаты этих исследований сведены в табл. 3.2.
Истинные гермафродиты. Исключительно редко встречающиеся индивиды, в организме которых присутствуют ткани как яичников, так и яичек, а их наружные половые органы часто представляют собой сочетание женских и мужских структур.
Псевдогермафродиты. Индивиды, чьи гонады соответствуют их хромосомному полу, однако чья внутренняя и наружная репродуктивная анатомия представляет собой сочетание мужских и женских структур.
Таблица 3.2. Сводная таблица некоторых примеров аномальной пренатальной половой дифференциации
Синдром Тернера
Хромосомный пол:45, XO
Гонадный пол:Отдельные волокна тканей яичников
Внутренние репродуктивные структуры:Матка и фаллопиевы трубы
Наружные половые органы:Нормальные женские
Способность к продолжению рода:Стерильны
Вторичные половые признаки:Недоразвиты, грудные железы отсутствуют
Гендерная идентичность:Женская
Синдром Кляйнфелтера
Хромосомный пол:47, XXY
Гонадный пол:Маленькие яички
Внутренние репродуктивные структуры:Нормальные мужские
Наружные половые органы:Маленькие пенис и мошонка
Способность к продолжению рода:Стерильны
Вторичные половые признаки:Некоторая феминизация вторичных признаков; могут иметь развитые грудные железы и округлые формы тела
Гендерная идентичность:Обычно мужская, хотя имеет более высокий, чем в норме, процент случаев гендерной неопределенности
Синдром андрогенной невосприимчивости
Хромосомный пол:46, XY
Гонадный пол:Неопустившиеся яички, гениталии и неглубокая вагина
Внутренние репродуктивные структуры:Отсутствие нормального состава как мужских, так и женских внутренних структур
Наружные половые органы:Нормальные женские
Способность к продолжению рода:Стерильны
Вторичные половые признаки:В период полового созревания имеет место развитие грудных желез и появление других признаков, но менструации не начинаются
Гендерная идентичность:Женская
Внутриутробно андрогенизированные женщины
Хромосомный пол:46, XX
Гонадный пол:Яичники
Внутренние репродуктивные структуры:Нормальные женские
Наружные половые органы:Амбивалентные (как правило, более напоминающие мужские, чем женские)
Способность к продолжению рода:Способны
Вторичные половые признаки:Нормальные женские (индивидам с дисфункциями надпочечниковых желез может назначаться кортизон с целью предотвращения маскулинизации)
Гендерная идентичность:Женская, однако имеет место глубокая неудовлетворенность своей женской гендерной идентичностью; склонность к традиционно мужским занятиям