Страница:
запрещенной зоны
(например, у InSb она составляет 0,18
эв)
.Полупроводник наносят в виде тонкого слоя на стеклянную или кварцевую подложку либо вырезают в виде тонкой пластинки из монокристалла. Слой (пластинку) снабжают двумя контактами (электродами). Подложку с фоточувствительным слоем (или пластинку) и электроды помещают в защитный корпус.
Важнейшие параметры Ф.: интегральная чувствительность (определяемая как отношение изменения напряжения на единицу мощности падающего излучения при номинальном значении напряжения питания) составляет 10 3–10 8 в/вт;порог чувствительности (величина минимального сигнала, регистрируемого Ф., отнесённая к единице полосы рабочих частот) достигает 10 -12 вт/гц 1 / 2постоянная времени (характеризующая инерционность Ф.) лежит в пределах 10 -3–10 -8 сек.Для повышения порога чувствительности и расширения рабочего диапазона длин волн принимаемого излучения фоточувствительный слой некоторых Ф. подвергают охлаждению. Так, охлаждение Ф. из PbS до 78 К позволяет на порядок повысить пороговую чувствительность и расширить диапазон длин волн принимаемого излучения с 3,3 мкмдо 5 мкм;глубоким охлаждением (до 4 К) Ф. из Ge, легированного Zn, доводят границу его спектральной чувствительности до 40 мкм.
Лит.:Марков М, Н., Приемники инфракрасного излучения, М., 1968; Аксененко М. Д., Красовский Е. А., Фоторезисторы, М., 1973.
И. Ф. Усольцев.
фото...
и
рецепторы
)
,световоспринимающие. светочувствительные образования, способные в ответ на поглощение квантов света молекулами содержащихся в них пигментов генерировать физиологический (нервный, рецепторный) сигнал. В широком смысле под Ф. понимают все светочувствительные образования от
стигмы
одноклеточных организмов и одиночных рассеянных по телу светочувствительных клеток (черви, ланцетник) до специализированных зрительных клеток глаза – сложного органа фоторецепции животных и человека. К Ф. относят также различные структуры – хлоропласты растений, пластиды водорослей, хроматофоры бактерий, содержащие пигменты и обеспечивающие фотобиологические процессы (фотосинтез, фототропизм, фототаксис, фотопериодизм и др.).
Важнейшие параметры Ф.: интегральная чувствительность (определяемая как отношение изменения напряжения на единицу мощности падающего излучения при номинальном значении напряжения питания) составляет 10 3–10 8 в/вт;порог чувствительности (величина минимального сигнала, регистрируемого Ф., отнесённая к единице полосы рабочих частот) достигает 10 -12 вт/гц 1 / 2постоянная времени (характеризующая инерционность Ф.) лежит в пределах 10 -3–10 -8 сек.Для повышения порога чувствительности и расширения рабочего диапазона длин волн принимаемого излучения фоточувствительный слой некоторых Ф. подвергают охлаждению. Так, охлаждение Ф. из PbS до 78 К позволяет на порядок повысить пороговую чувствительность и расширить диапазон длин волн принимаемого излучения с 3,3 мкмдо 5 мкм;глубоким охлаждением (до 4 К) Ф. из Ge, легированного Zn, доводят границу его спектральной чувствительности до 40 мкм.
Лит.:Марков М, Н., Приемники инфракрасного излучения, М., 1968; Аксененко М. Д., Красовский Е. А., Фоторезисторы, М., 1973.
И. Ф. Усольцев.
 и диаметром от 1–2 до 6–8
мкм(в зависимости от вида животного); диски ориентированы строго перпендикулярно длинной оси клетки: в палочках они «плавают» в цитоплазме, т.к. оторваны от наружной клеточной мембраны, в большинстве же колбочек они сохраняют с ней связь. В палочках, но не в колбочках происходит постоянное обновление наружного сегмента за счёт образования новых и отмирания (фагоцитоза) «старых» верхушечных дисков. Вследствие строгой ориентации молекул зрительного пигмента в фоторецепторной мембране и особой (трубчатой) упаковки её в клетке многие беспозвоночные способны различать направление поляризации света и ориентироваться по нему. Палочки позвоночных – рецепторы сумеречного (скотопического) зрения, колбочки ответственны за дневное (фотопическое) и
цветовое зрение.
Фасеточные глазанасекомых также способны к различению цвета.
и диаметром от 1–2 до 6–8
мкм(в зависимости от вида животного); диски ориентированы строго перпендикулярно длинной оси клетки: в палочках они «плавают» в цитоплазме, т.к. оторваны от наружной клеточной мембраны, в большинстве же колбочек они сохраняют с ней связь. В палочках, но не в колбочках происходит постоянное обновление наружного сегмента за счёт образования новых и отмирания (фагоцитоза) «старых» верхушечных дисков. Вследствие строгой ориентации молекул зрительного пигмента в фоторецепторной мембране и особой (трубчатой) упаковки её в клетке многие беспозвоночные способны различать направление поляризации света и ориентироваться по нему. Палочки позвоночных – рецепторы сумеречного (скотопического) зрения, колбочки ответственны за дневное (фотопическое) и
цветовое зрение.
Фасеточные глазанасекомых также способны к различению цвета.
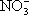 ) и сульфаты (
) и сульфаты (
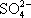 ). Фотосинтезирующие бактерии не выделяют и не используют молекулярный кислород (большинство из них облигатные, т. е. обязательные
анаэробы
)
.Вместо воды в качестве доноров электронов эти бактерии используют либо неорганические соединения (сероводород, тиосульфат, газообразный водород), либо органические вещества (молочную кислоту, изопропиловый спирт). Источником углерода в большинстве случаев является также CO
2, но наряду с этим и некоторые органические соединения (например, ацетат). Т. о., Ф. у разных организмов может протекать с использованием различных доноров (ДН
2), акцепторов (А) электронов и водорода и может быть представлен схематически обобщённым уравнением:
). Фотосинтезирующие бактерии не выделяют и не используют молекулярный кислород (большинство из них облигатные, т. е. обязательные
анаэробы
)
.Вместо воды в качестве доноров электронов эти бактерии используют либо неорганические соединения (сероводород, тиосульфат, газообразный водород), либо органические вещества (молочную кислоту, изопропиловый спирт). Источником углерода в большинстве случаев является также CO
2, но наряду с этим и некоторые органические соединения (например, ацетат). Т. о., Ф. у разных организмов может протекать с использованием различных доноров (ДН
2), акцепторов (А) электронов и водорода и может быть представлен схематически обобщённым уравнением: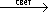 AH
2+ Д,
AH
2+ Д,